
As plantas precisam de elementos químicos essenciais da Tabela Periódica para crescer, se desenvolver e realizar suas funções vitais. Esses nutrientes se dividem em macronutrientes (necessários em grandes quantidades) e micronutrientes (necessários em pequenas quantidades).
A fotossíntese é um processo pelo qual as plantas, seres autotróficos, transformam energia luminosa em energia química processando o dióxido de carbono (CO2), água (H2O) e minerais em compostos orgânicos e oxigênio gasoso (O2) (Figura 1).
Como as plantas se “alimentam” dos elementos químicos da tabela periódica, que são essenciais as suas vidas e, na sequência, para também para os outros seres vivos
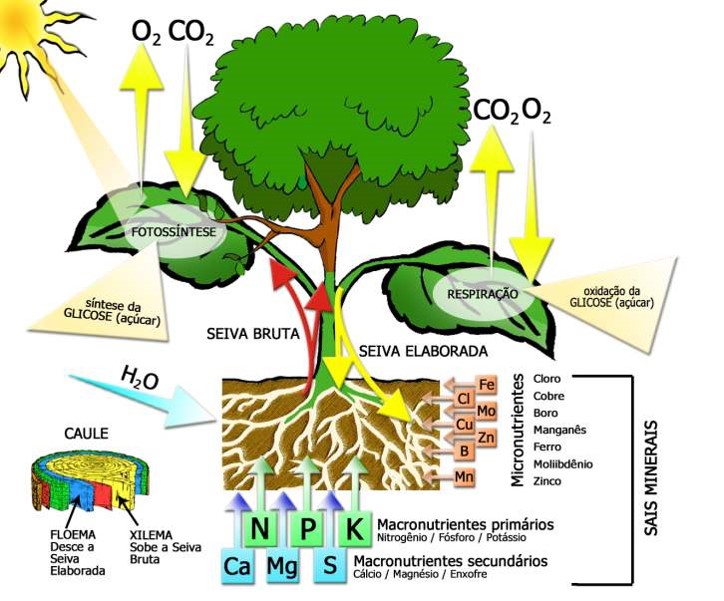
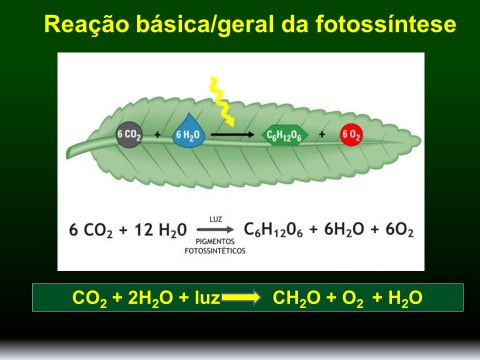
Figura 1. Equação geral da fotossíntese.
Pela fotossíntese as plantas produzem o seu próprio alimento, constituído essencialmente por açúcares, como a glicose. A fotossíntese dá início a toda a cadeia alimentar. Sem esta, os animais e os outros seres heterotróficos seriam incapazes de sobreviver, já que a base da sua alimentação está nas substâncias orgânicas proporcionadas pelas plantas verdes. É por meio dela que os vegetais produzem alimentos, combustível indispensável para a vida da planta, do homem e de outros animais.
As folhas possuem células denominadas fotossintetizadoras, que contém clorofila e são muito sensíveis à luz. Quando a luz incide em uma molécula de clorofila, esta absorve parte da energia luminosa que permite a reação do gás carbônico com água, produzindo carboidratos e liberando oxigênio.
A água é retirada do solo pela raiz e sobe como seiva pelos vasos. O gás carbônico é retirado da atmosfera e absorvido pelas folhas. A energia luminosa vem da luz solar. A clorofila tem ação catalisadora na reação. Isto é, não se desgasta nem é consumida, apenas ativa a reação.
A absorção da energia luminosa e sua transformação em energia química permitem o crescimento das plantas, seu florescimento e a produção de frutos.
A luz utilizada nessa formação é absorvida por uma série de pigmentos. Cada pigmento absorve determinados comprimentos de ondas, refletindo os que não absorve. A cor do pigmento é dada pelo comprimento de onda refletido, podendo-se determinar o espectro de absorção de cada pigmento através de um espectrofotômetro.
A fotossíntese é um processo redox em que a água é oxidada e dióxido de carbono é reduzido. A liberação do oxigênio da fotossíntese provém da água e não do dióxido de carbono (Figura 2).
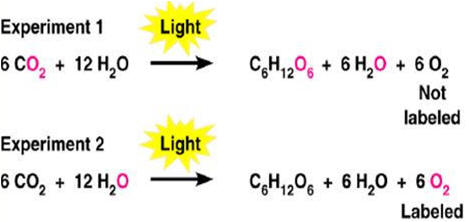
Figura 2. Carbono marcado mostra que o oxigênio liberado na fotossíntese provém da água.
O ângulo de inserção foliar regula o grau de penetração no dossel. Os arranjos mais erectófilos apresentam menor coeficiente de extinção, resultando numa distribuição da radiação incidente por maior área foliar, proporcionando assim, maior taxa de fotossíntese.
Alguns autores consideram que o ângulo das folhas não afeta somente a iluminação relativa das folhas totalmente expostas, mas também a área de sombra projetada pelas folhas e, conseqüentemente, o fluxo de luz disponível para as folhas inferiores. Levando-se em conta que as respostas das taxas fotossintéticas à intensidade de iluminação são curvilíneas, pode-se afirmar que maiores produções e utilização mais eficiente da luz são conseguidas iluminando-se muitas folhas a níveis moderados de intensidade luminosa do que expondo algumas poucas folhas a iluminação intensa.
As diferenças de interceptação de luz e produção fotossintética relacionadas com o ângulo foliar ocorrem principalmente para a luz direta e, são também dependentes da elevação solar. Para o ângulo foliar de 0 ºC (folhas planas) são pequenas as variações que ocorrem na produção de matéria seca em função do IAF ou da hora do dia. Já para o ângulo de 80 ºC, a produção é maior quanto maior for o IAF (até certos limites), ocorrendo diferenças principalmente nas horas de maior disponibilidade de radiação direta, sendo a incidência desta radiação bastante vertical.
Um pouco da Evolução histórica dos conhecimentos sobre a fotossíntese
Na Grécia antiga acreditava-se que as plantas obtinham do solo e da água todos os elementos necessários ao seu crescimento. No século 17, o médico Van Helmont cultivou uma planta em um jarro com terra, regando a planta somente com água da chuva. Ele observou bom crescimento da planta após 5 anos, mas a quantidade de terra no jarro quase não decresceu. Van Helmont concluiu que o material utilizado pela planta para o seu crescimento veio da água utilizada para regá-la. Em 1727, o botânico inglês Stephan Hales observou que as plantas usavam principalmente o ar como fonte de nutrientes para o seu crescimento (Calvin, 1961)- Século XVII: Jan Baptista van Helmont (1579-1644).
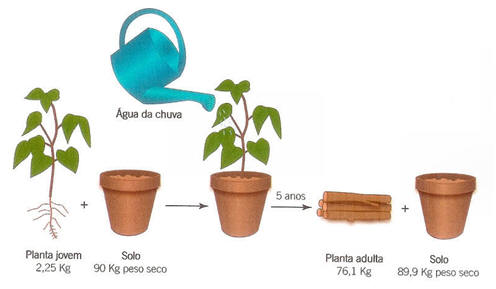
Figura 3. Experimento realizado por Jan Baptista van Helmont (fonte: PRISCO, 1989).
Foi somente no século XVIII, mais precisamente em 1727, que Stephan Hales sugeriu que parte da nutrição da planta dependia da atmosfera, tendo a luz papel importante neste processo. Nesta época, ainda não se conhecia a composição química do ar e nem se tinha ideia de como acontecia à respiração dos animais. Os alquimistas, tentando explicar o fenômeno da combustão, criaram a teoria de que quando, por exemplo, uma vela queimava havia a produção de uma “substância tóxica”, denominada flogisto (fluido produzido como resultado da combustão), que fazia com que o ar se tornasse impuro ou contaminado (Prisco, 1989).
Em 1771, o inglês Joseph Priestley descobriu que se um rato era colocado sob uma campânula juntamente com uma vela acesa, depois de algum tempo o animal morria. Sua interpretação foi que o ar estava contaminado devido a combustão da vela, a qual produzia “flogisto”. Quando ele substituiu o rato por uma planta, ela se desenvolveu normalmente. Isto foi interpretado por ele como sendo devido à capacidade que têm as plantas de purificar o ar, ou seja, de “desflogistá-lo”.

Figura 4. Joseph Priestly mostrou que o ar que fora emitido pela vela e pelo camundongo poderia ser restaurado por uma planta.
Ao tomar conhecimento das experiências de Priestley, o cientista holandês Jan Ingen-Housz deu continuidade ao trabalho e em 1779 concluiu que a “purificação do ar” feita pelas plantas dependia da luz e que isto só ocorria nas partes verdes da planta. As partes não verdes (raízes, por exemplo) comportavam-se de maneira idêntica aos animais. Nesta época, o químico francês Antoine Lavoisier esclareceu o fenômeno da combustão, demonstrando que neste processo o que ocorre é o consumo de oxigênio com conseqüente liberação de gás carbônico, colocando por terra a teoria do flogisto. De posse desta informação, Ingen-Housz e o suíço Jean Senebier (1782) concluíram que o CO2 existente no ar era a fonte de carbono para a formação da matéria orgânica vegetal. As experiências até aqui relatadas eram qualitativas, mas o suíço Nicholas de Saussure (1804) deu um cunho mais quantitativo aos seus experimentos, podendo, assim, chegar a conclusão de que a água era também um reagente da fotossíntese. Além disto, ele demonstrou claramente que na presença de luz as plantas absorviam CO2 e liberavam O2 e que no escuro acontecia o inverso (Prisco, 1989). Durante o restante do século XIX as contribuições dos alemães Julius Robert Meyer (1842) e Julius von Sachs (1864) permitiram entender a fotossíntese, não só como um processo de trocas gasosas mas, também, como um processo em que há síntese de matéria orgânica e transformação de energia luminosa em energia química (Prisco, 1989).
Em 1905, o fisiologista inglês F. F. Blackman, estudando os efeitos da temperatura, da concentração de CO2 e da intensidade luminosa sobre a fotossíntese, chegou à importante conclusão de que este processo consistia de dois tipos de reações: as que dependiam da luz e aquelas que ocorriam no escuro. As reações da luz eram rápidas e a temperatura não as afetava, já as reações do escuro eram lentas e dependiam da temperatura, ou seja, as reações da luz eram fotoquímicas e as do escuro eram bioquímicas (Prisco, 1989).
Durante a década de 1920, o microbiologista holandês C. B. van Niel observou que existiam bactérias que eram capazes de fotossintetizar, mas que não liberavam O2 durante este processo. Ele observou também que estes microorganismos, ao invés de H2O usavam H2S como reagente da fotossíntese (Prisco, 1989).
CO2 + 2H2S +Luz → (CH2O) + H2O + 2S
A comparação da equação acima com a da fotossíntese de plantas verdes o levou a concluir que H2O e H2S desempenham papel semelhante, isto é, são doadores de hidrogênio.
Portanto, a equação geral da fotossíntese pode ser escrita como:
CO2 + 2H2A + Luz → (CH2O) + H2O + 2A (agente promotor reação: Organismos Fotossintetizantes)
Além disso, ele postulou que o O2 liberado na fotossíntese provém da água e não do CO2, como se imaginava na época. Foi também este cientista holandês que lançou a idéia de que a luz é que produz o agente redutor (H) e o agente oxidante (oxigênio) era produzido a partir da água, processo que ele denominou de fotólise da água (Prisco, 1989).
Niel concluiu que H2O e H2S desempenham papel semelhante, isto é, são doadores de hidrogênio. Além disso, ele postulou que o O2 liberado na fotossíntese provém da água e não do CO2, como se imaginava na época. Foi também este cientista holandês que lançou a idéia de que a luz é que produz o agente redutor (H) e o agente oxidante (oxigênio) era produzido a partir da água, processo que ele denominou de fotólise da água (Prisco, 1989).
O bioquímico inglês Robert Hill (1937) demonstrou que preparações contendo fragmentos de folhas ou cloroplastos isolados, na presença de água, luz e de um aceptor artificial de elétrons ou de hidrogênio (oxalato férrico, cianeto férrico ou ferricianeto de potássio) podiam provocar a liberação de oxigênio (Prisco, 1989).
Após a segunda guerra mundial, ocorreram inúmeras descobertas importantes para a elucidação do processo fotossintético. A primeira delas, ocorrida na década de 1950, foi a demonstração feita pela bioquímica americana Mary Allen, de que preparações de cloroplastos eram capazes de fixar CO2 na presença de luz e de água, ou seja, ela provou experimentalmente o que Hill havia postulado em 1937. Foi também na mesma época que outro americano, Daniel Arnon, demonstrou que o sistema de membranas de cloroplastos isolados era capaz de sintetizar ATP e NADPH, na presença de luz. Após esta série de descobertas pôde-se concluir que durante as reações da luz há liberação de O2, produção de ATP (energia) e NADPH (poder redutor) e que estas reações ocorriam no sistema de membranas dos cloroplastos (Prisco, 1989).
FOTOSSÍNTESE (Metabolismo Fotossintético)
(da absorção da radiação, utilização dos elétrons e prótons da agua e da fixação e redução do dióxido de carbono pelas folhas, até a produção de trioses pelo ciclo de CALVIN nos cloroplastos e sacarose no citossol).
Para um melhor entendimento e compreensão de todo o evento fotossintético e seus possíveis pontos e aspectos de sua regulação e funcionamento sugere-se, que num primeiro momento, o estudo da fotossintese pode ser, didaticamente, em três grupos de processos:
- Processo Fotoquímico (depende diretamente da radiação solar-natural, ocorre nos TILACÓIDES, produz ATP, NADPH e O2 ): Radiação fotossintéticamente ativa RFA/PAR; Enverdecimento de plantas: ontogênese de cloroplastos e biossíntese de pigmentos; Fotossistemas (PSI/PSII) e a produção de ATP e de NADPH; Fatores que regulam e interferem no processo fotoquímico, Inibidores da fotoquímica;
- Processo de Difusão de CO2: abordagem das peculiaridades e detalhes de como o dióxido de carbono vai dos vários pontos da micro e macro atmosfera ate o local onde se encontra a enzima fixadora RuBisCO e/ou PEPC/PEPCase em plantas C4 e CAM (citossol) e plantas C3 (estroma do cloroplasto); Fatores que regulam e interferem no processo de difusão do CO2 da atmosfera até o local de fixação, em plantas C3, C4 e CAM
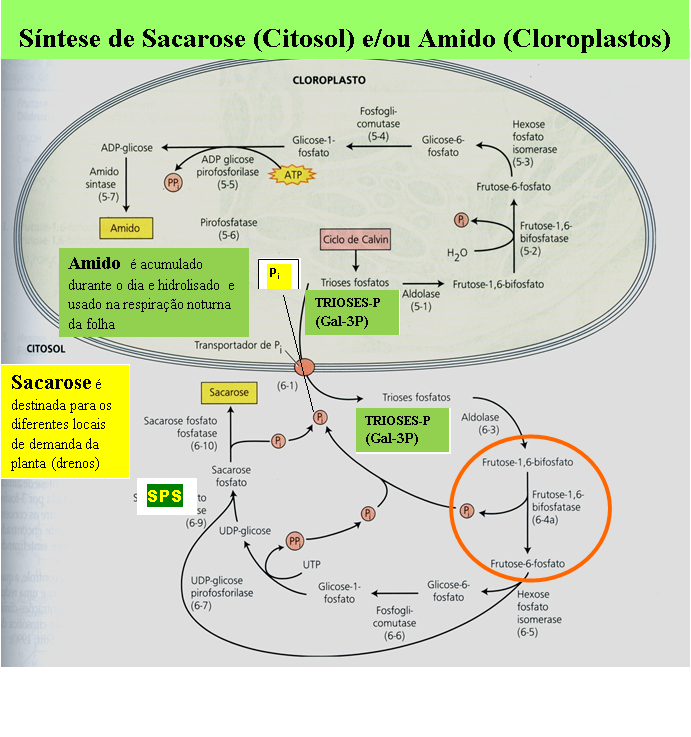
- PROCESSO FIXAÇÃO E REDUÇÃO DO CO2: a enzima fixadora RuBisCO está no estroma das células do mesofilo nas plantas C3 e no estroma das células da bainha; a enzima PEPC/PEPCase em plantas C4 e CAM está no citossol, sempre nas células do mesofilo. A redução do carbono e a produção de TRIOSES em todos os tipos de plantas ocorre sempre no estroma, produzindo amido no interior do Cloroplasto e SACAROSE no citossol que vai ser exportada, via floema, para atender a demanda de outros órgãos. O ATP e NADPH produzidos pelo processo fotoquimico são utilizados para reduzir o CO2 e produzir as trioses
- Fatores edafoclimáticos e da própria planta que regulam os Processos envolvidos na fotossíntese (ponto de saturação de luz e de compensação de luz e de CO2); Agua, CO2, radiação, temperatur, IAF,TAL, etc
PRODUÇÃO DAS TRIOSES PELA FOTOSSÍNTESE E EM SEGUIDA A SUBSEQUENTE PRODUÇÃO DAS HEXOSES PARA A SINTESE DE AMIDO NO ESTROMA DO CLOROPLASTOS E DE SACAROSE NO CITOSOL DAS CELULAS FOLIARES QUE VAI SER EXPORTADA PARA ATENDER A DEMANDA DE TODOS OS DEMAIS ÓRGÃOS DA PLANTA
Algumas das principais bibliografias históricas sobre fotossíntese que vale a pena conhece
1. ARONOFF, S. AND L. VERNON. 1950. Metabolism of soybean leaves. I. The sequence of formation of the soluble carbohydrates during photosynthesis.Arch. Biochem. 28: 424-39.
2. BADIN, E. J. AND M. CALVIN. 1950. Photoreduction and the hydrogen-oxygen-carbon dioxide dark reaction. J. Am. Chem. Soc. 72: 5266-70.
3. BASSHAM, J. A., A. A. BENSON, L. D. KAY, A. Z. HARRIs, A. T. WILSON, AND M. CALVIN. 1954. The cyclic regeneration of carbon dioxide acceptor. J. Am. Chem. Soc. 76: 1760-70.
4. BASSHAM, J. A., A. A. BENSON, AND M. CALVIN. 1950. The role of malic acid. J. Biol. Chem. 185: 781-87.
5. BENSON, A. A., S. KAWAUCIHI, P. HAYES, AND M. CALVIN. 1952. The path of carbon in photosynthesis. J. Am. Chem. Soc. 74: 4477-82.
6. BONDE, E. K. 1952. The influence of carbon dioxide concentration upon the rate of photosynthesis in Sinapis alba. Physiol. Plantarum 5: 298-304.
7. BONNET, J. A. 1962. Chemical concept of sucrose formation and maturity status of harvested sugarcane in Puerto Rico. Sugar J. 25: 45-54.
8. BURR, G. 0. 1962. The use of radioisotopes by the Hawaiian sugar plantations. Intern. J. Appl. Radiation Isotopes. 13: 365-74.
9. CALVIN, M., J. A. BASSHAM, AND A. A. BENSON. 1950. Chemical transformations of carbon in photosynthesis. Federation Proc. 9: 524-34.
10. HARTr, C. E. 1963. Translocation of sugar in the cane plant. Hawaiian Sugar Technol. Rep. 22:151-67.
11. KORTSCHAK, H. P., C. E. HARTr, AND G. 0. BURR. 1957. PGA and photosynthesis in sugar cane. Proc. Hawaiian Acad. Sci. p 21.
12. LEDON, A. C. AND F. A. Z. GONZALES. 1950. In-dustrialization of photosynthesis through the use of sugar cane. Proc. Cuban Sugar Technol. 24:563-72.
13. MANGELSDORF, A. J. 1953. Sugar cane breeding in Hawaii. Hawaiian Planters’ Record 54(3):101-37.
14. SEIDELL, A. 1941. Solubilities of organic compounds. 3rd ed. Van Nostrand and Company, p232.
15. SINGH, N. AND K. N. LAL. 1935. Limiting factors and photosynthesis. Plant Physiol. 10: 245-68.