
A fotossíntese C4 é uma via fotossintética adaptativa que evoluiu para diminuir os efeitos prejudiciais do declínio gradual de CO2 atmosférico, como a fotorrespiração, sendo conhecida também como via de Hatch-Slack. É comum que em plantas C4, dois tipos de células cooperem no processo de fixação de carbono: da bainha e do mesofilo. Embora grande parte da bioquímica está bem caracterizada, pouco se sabe sobre os mecanismos genéticos subjacentes à especificidade de células do tipo de condução C4. No entanto vários estudos têm mostrado que age na regulação em múltiplos níveis, incluindo transcricional, pós-transcricional, pós-traducionais e epigenéticos (MARENCO & LOPES, 2009).
Esse ciclo é típico de gramíneas tropicais, cana-de-açúcar e milho, além de ocorrer em 16 famílias tanto de monocotiledôneas quanto de dicotiledôneas e é particularmente proeminente nas famílias Poaceae (milho, milheto, sorgo, cana-de-açúcar), Ciperaceae e Chenopodiaceae. Nessa última, o gênero Atriplex possui espécies C3 e C4, o que permite uma comparação mais próxima entre as duas vias fotossintéticas para estudos sobre o comportamento de plantas C4 (BJÖRKMAN et al., 1971; PIMENTEL, 1985). Aproximadamente 1% das espécies conhecidas possuem metabolismo C4. É importante ressaltar também que oito entre as dez ervas invasoras mais importantes na agricultura usam a via C4 (NOBEL,1991).
A anatomia “Kranz” (coroa ou grinalda em alemão) já era conhecida por HABERLANDT (1884), sem que se soubesse o seu significado fisiológico. As plantas C4 possuem a anatomia do tipo “Kranz”, com células distintas no mesofilo e na bainha perivascular. Entretanto, durante esse processo evolutivo pode-se observar estágios intermediários em que as células não possuem uma caracterização completa, nem da via C3, nem da via C4. Essas espécies são chamadas de intermediárias C3-C4. Nesse processo pode-se observar modificações graduais tanto anatômicas, quanto bioquímicas, até o total estabelecimento do ciclo C4.
Nas plantas C4, a parede celular, entre as células externas da bainha perivascular e as células mais internas do mesofilo, possui um número de plasmosdemos muito superior às outras paredes celulares, permitindo assim o trânsito dos ácidos orgânicos para o metabolismo C4, e também o trânsito de trioses-P, que transportarão energia e poder redutor.
Na via C4, o CO2 atmosférico é fixado em um ácido orgânico com 3C, o fosfoenolpiruvato (PEP) nas células do mesofilo (onde há a atuação da enzima PEP carboxilase- PEPcase), produzindo ácidos orgânicos de 4C (Figura 1). Tais ácidos orgânicos de 4C migram para a bainha perivascular (onde haverá a atuação da enzima Rubisco), onde são descarboxilados, liberando CO2, e um ácido orgânico de 3C, que retorna ao mesófilo para regeneração do substrato de 3C, visando a carboxilação primária. Portanto, na via C4 não ocorre produção de carboidratos, e ela servirá somente para aumentar a concentração do CO2 na bainha perivascular, favorecendo a ação carboxilase da rubisco (PEISKER & HENDERSON, 1992).
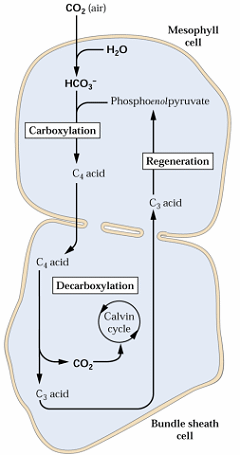 Figura 1. Esquema simplificado da via fotossintética C4.
Figura 1. Esquema simplificado da via fotossintética C4.
Essa via C4 inicia-se, então, pela fixação do CO2 no PEP (3C), formando o ácido oxaloacético com 4C, pela ação de fosfoenolpiruvato carboxilase (PEP-case) que se encontra no citoplasma. O ácido oxaloacético produzido pode, então, ser transformado em ácido málico (MAL), com gasto de 1 NADPH, ou em ácido aspártico (ASP). Estes ácidos orgânicos com 4C, o málico ou o aspártico, migram então para a bainha perivascular, onde serão descarboxilados por uma das três enzimas de descarboxilação, liberando CO2 para ser incorporado à RuBP, no ciclo de Benson-Calvin. De acordo com a enzima de descarboxilação do ácido com 4C, as plantas serão classificadas em três tipos. Os ácidos com 3C resultantes da descarboxilação, o ácido pirúvico (PIR) ou a alanina (ALA), retornam ao mesófilo consumindo 2 a 3ATP para regenerar o substrato da carboxilação primária C4, o ácido fosfoenolpirúvico (RICHTER, 1993). Portanto, a via C4 serve para transportar e concentrar o CO2 na bainha perivascular, onde se dará a sua redução a açúcares pela via C3, que é a única via metabólica de síntese de carboidratos.
Essa concentração de CO2 na bainha perivascular aumentará a eficiência de síntese de açúcares, pois com alto teor de CO2 haverá maior atividade carboxilase da rubisco, em detrimento da atividade oxigenase. Há um gasto em torno de 5 ATP e 2 NADPH na via C4, porém o aumento do fluxo de H+ via ciclo Q nas plantas C4, produzirá ATP complementar (FURBANK et al, 1990). A regulação dos eventos C4 no mesófilo já está bem caracterizada, porém a regulação da descarboxilação e o transporte de metabólitos na bainha perivascular são menos esclarecidos, sobretudo pela dificuldade de se conseguir grandes quantidades de tecido dessa bainha. Os estudos sobre o controle da fotossíntese C4 estão ainda no começo, mas os trabalhos que estão sendo feitos com mutantes de Amaranthus edulis , sem algumas enzimas C4, e plantas transgênicas de Faveria bidensis, no CSIRO-Austrália, poderão impulsionar o conhecimento dos mecanismos de controle da fotossíntese C4 (LEEGOOD, 1996).
Os ácidos C4 malato e aspartato são os primeiros intermediários estáveis detectáveis da fotossíntese em folhas de cana-de-açúcar e o átomo de carbono 4 do malato, consequentemente, torna-se o carbono 1 do 3-fosfoglicerato. A carboxilação inicial nessas folhas não é catalisada pela rubisco, mas pela PEP (fosfoenolpiruvato) carboxilase.
A maneira como o carbono é transferido da posição 4 do malato para a posição 1 do 3-fosfoglicerato tornou-se clara com a elucidação do envolvimento das células do mesofilo e da bainha vascular. As enzimas que participam do processo ocorrem em um dos dois tipos de células – PEP carboxilase e a piruvato ortofosfato diquinase – são restritas às células do mesofilo; as descarboxilases e as enzimas do ciclo completo de Calvin estão confinadas à célula da bainha vascular.
A regeneração do aceptor primário – fosfoenolpiruvato – consome duas ligações fosfato de alta energia: uma na reação catalisada pela piruvato ortofosfato diquinase e outra na conversão do PP a 2Pi, catalisada pela pirofosfatase.
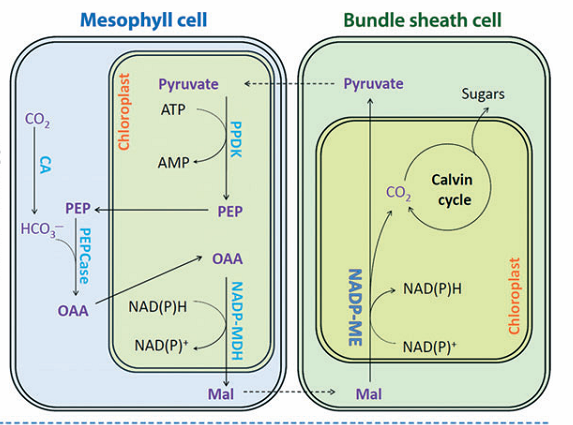
Figura 2. Esquema da via C4 com indicação de seus intermediários. Mal- malato; CA- anidrase carbônica; NADP-MDH- Malato desidrogenase dependente de NADP; PPDK- Piruvato fosfato diquinase; NADP-ME- Enzima málica dependente de NADP.
O transporte pelos plasmodesmas entre as células do mesofilo e da bainha vascular é acionado por gradientes de difusão. O transporte dentro das células é regulado por gradientes de concentração e pela operação de transportadores especializados no envoltório do cloroplasto. Assim, o ciclo transfere efetivamente CO2 da atmosfera para as células da bainha vascular. Esse processo de transporte gera uma concentração de CO2 nas células na bainha vascular muito mais elevada do que poderia ocorrer se houvesse equilíbrio com a atmosfera externa da folha. Esta concentração elevada de CO2 nos sítios de carboxilação da RuBisCO resulta na supressão da oxigenação da ribulose-1,5-bifosfato e, por conseqüência, da fotorrespiração.
Além das diferenças bioquímicas existentes entre esses dois tipos celulares, pode-se perceber também diferenças entre o aparato fotoquímico. Os cloroplastos do mesofilo possuem grande quantidade de tilacóides empilhados formando os granas, que permite melhor capacidade de funcionamento dos fotossistemas. Já os cloroplastos da bainha, não apresentam o empilhamento dos granas, enquanto em seu estroma há uma grande quantidade de grânulos de amido, evidenciando sua atividade fotossintética mais voltada à etapa bioquímica. Desta forma, em plantas C4, o processo fotossintético se divide basicamente da seguinte forma: os cloroplastos da célula do mesofilo são responsáveis pela etapa fotoquímica, enquanto os da bainha do feixe pela etapa bioquímica (Figura 3).
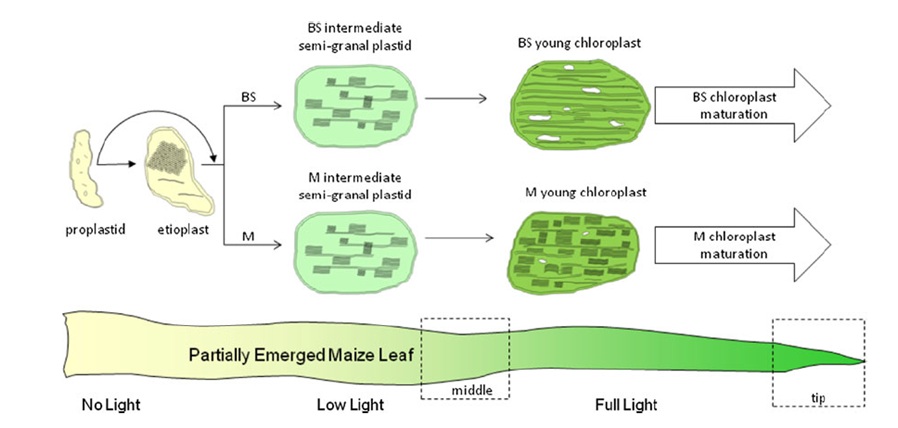 Figura 3. Esquema mostrando o processo de diferenciação de cloroplastos (em folhas de milho) a partir de etioplastos, gerando cloroplasto das células do mesofilo e da bainha (Sharpe et al., 2011).
Figura 3. Esquema mostrando o processo de diferenciação de cloroplastos (em folhas de milho) a partir de etioplastos, gerando cloroplasto das células do mesofilo e da bainha (Sharpe et al., 2011).