
Os principais locais na planta para a redução do NO3– são folhas e raízes. A maioria das espécies já estudadas apresentam atividade da enzima redutase do nitrato (RN) nas folhas. Entretanto, a importância relativa da raiz e folha na assimilação do NO3– depende de dois fatores: a atividade da RN na raiz e a disponibilidade de NO3– no meio. Espécies com capacidade muito baixa em assimilar o NO3– nas raízes (por exemplo, espécies de Gossypium, Xanthium e Cucumis) enviam todo o íon absorvido (via xilema) para assimilação nas folhas. Espécies com alta capacidade em assimilar o NO3– nas raízes (por exemplo, Lupinus spp.) dificilmente têm essa capacidade superada pelo NO3– absorvido, e, consequentemente, a importância da folha é pequena. Porém, a maioria das espécies são intermediárias em termos de capacidade de assimilar o NO3– nas raízes. Nesses casos, a folha toma-se importante apenas quando o NO3– no meio estiver em concentração suficiente para superar a capacidade de redução da raiz. No entanto, há exceções a essa regra. Apesar de uma capacidade razoável para a assimilação do NO3– na raiz, algumas leguminosas transportam parte significativa do NO3– para a folha mesmo quando a capacidade da raiz não é superada.
Na célula a redução ocorre em duas etapas, cada uma envolvendo doadores de elétrons específicos:
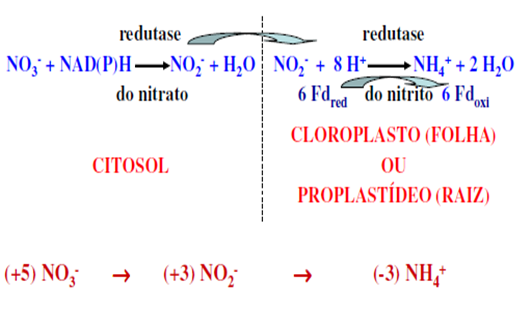
O primeiro passo é catalisado pela enzima redutase do nitrato (RN), localizada no citoplasma, enquanto a redutase do nitrito (RNi), localizada no cloroplasto (tecidos verdes) ou plastídeos (tecidos não-verdes), catalisa o segundo. Na maioria das espécies estudadas, a enzima RN tem NADH como doador específico de elétrons. Entretanto, em algumas espécies a enzima utiliza tanto NADH como NADPH. Essa enzima bi-específica pode ocorrer isoladamente ou junto com a enzima monoespecífica. Em soja são conhecidas três isoformas, uma induzida pelo NO3– e específica para NADH e duas formas constitutivas, uma específica para NADH e outra bi-específica. A estrutura molecular da RN é bastante complexa, sendo a enzima constituída de duas subunidades idênticas de 110 a 115 kDa. Cada subunidade é composta de regiões, distintas, envolvidas na transferência de elétrons do NADH até o NO3–.
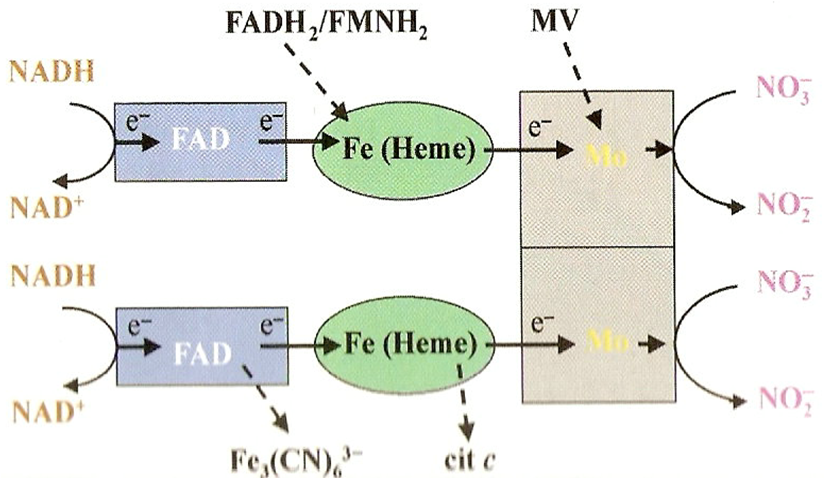
A presença de molibdênio na proteína como co-fator é interessante pelo fato de a RN ser uma das poucas proteínas conhecidas em plantas que contêm esse íon. Na deficiência de molibdênio, a atividade de RN fica bastante reduzida. Além do NO3– a RN pode transformar o clorato em cloreto, que é bastante tóxico para as plantas. Essa característica é explorada em herbicidas à base de clorato.
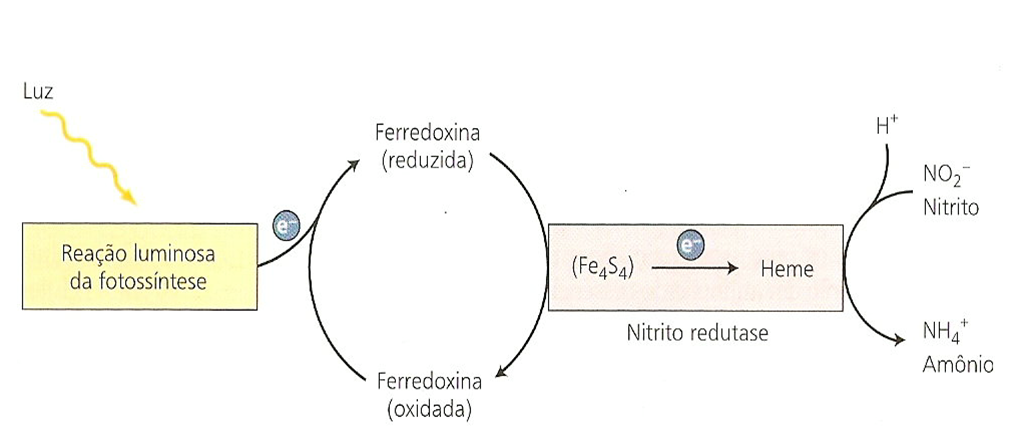
A redutase do nitrito (RNi) reduz o nitrito a amônia, onde o Fd representa a ferredoxina que pode estar reduzida ou oxidada. A Fd reduzida deriva do transporte de elétrons da fotossíntese nos cloroplastos, e do NADPH gerado pela via da oxidação da pentose fosfato nos tecidos não clorofilados.
Em função da importância estratégica da RN no metabolismo do nitrogênio em plantas (pois constitui a principal porta de entrada do N no metabolismo da planta), é natural que existam vários mecanismos de controle da sua atividade.
A eficiência do processo de assimilação do NO3– é maior na folha. Na raiz ou em outros tecidos não-verdes, a redução do NO3– e assimilação de NH4+ dependem de energia química do metabolismo de fotoassimilados fornecidos pelas folhas. Dessa forma consomem energia fotoquímica utilizada na fixação do gás carbônico. Na folha a necessidade de gasto de energia fotoquímica para a assimilação do nitrato é diferenciada em plantas tipo C3 e C4. No cloroplasto, isso nem sempre acontece, pois, os seis elétrons utilizados na redução do nitrito podem ser fornecidos diretamente pelas reações fotoquímicas, sem que haja competição com a fixação do gás carbônico. Pelo menos isso é possível sob alta intensidade luminosa, quando há excesso de energia fotoquímica e a assimilação do carbono satura facilmente.