
Para um melhor entendimento de como os íons são absorvidos pelo sistema radicular, é importante o conhecimento de algumas características e propriedades da parede celular e da membrana plasmática. É importante ressaltar que nas celulas os diferentes compartimentos são delimitados por membranas que possuem propriedades especificas relacionadas tambem com o movimento intracelular de ions minerais e biomoleculas orgânicas e com eventos importantes importantes como a fotoquimica da fotossintese nos cloroplatideos e cadeia de transporte de elétrons na respiração aeróbica nas mitocondrias.
1 – Parede Celular
A parede celular é um fino envoltório celulósico, de aproximadamente 1μm de espessura, que envolve e protege o protoplasto. Suas principais funções são:
– Dar resistência mecânica a célula;
– Controlar o crescimento celular;
– Formar uma barreira mecânica a penetração de certos microorganismos patogênicos;
– Regular certas funções celulares.
A célula vegetal madura é constituída por 3 camadas distintas:
– Parede primária: é a primeira a se formar e é constituída principalmente de celulose, hemicelulose e substância pécticas;
– Parede secundária: difere da parede primária por faltar substâncias pécticas e por conter maior porcentagem de celulose (mais força e rigidez). Pode conter lignina o que torna as células mas rígidas e impede o crescimento celular;
– Lamela média: localiza-se entre as células. Constitui um material amorfo, constituído de substâncias pécticas (grande quantidade de Ca). Tem a função de manter as células coesas e cimentadas.
Grande número de orifícios (pontuações) ocorrem na parede secundária em várias disposições e tamanhos (ficando a parede celular resumida à parede primária), conectando as células adjacentes. Esta conexão é feita por plasmodesmas que interconectam as células permitindo a troca de elementos minerais e outros materiais (Figura 1).
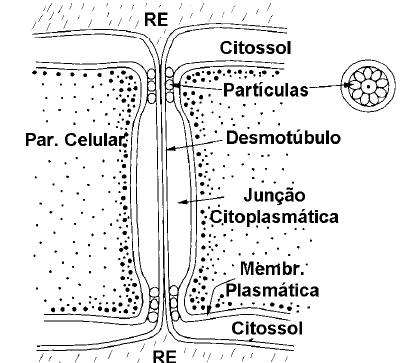
Figura 1: Diagrama da ultraestrutura de um plasmodesma. (Bonato, C. et al., 1998)
Os elementos minerais para atingir a membrana plasmática, primeiramente devem passar pela parede celular. A difusão através da parede celular depende:
- Espaço molecular
Há entre as microfibrilas e entre as micelas espaços, chamados interfibrilares, que possuem diâmetro médio de 10 e 1 nm, respectivamente. O tamanho destes espaços podem ser reduzido, até certo ponto, pela presença de matriz amorfa. Mesmo assim, a água e os pequenos solutos possuem diâmetro inferior a estes espaços, portanto, não há impedimento físico para os elementos minerais.
- Cargas elétricas
2 – Membrana Plasmática
A membrana plasmática é uma estrutura lipoprotéica extremamente fina (8 a 10 nm) que separa o protoplasto do meio exterior. Constitui uma efetiva barreira à livre movimentação de solutos, controlando o fluxo para dentro e para fora das células. Entretanto, não é uma barreira meramente passiva. Possui vários tipos de proteínas especializadas na promoção ou catálise de vários eventos metabólicos. São providas de bombas, transdutores de energia, receptores na sua superfície, logo, controlam a composição quantitativa e qualitativa do citoplasma influenciando profundamente no metabolismo celular.
As funções básicas principais são:
- Permeabilidade Seletiva
A membrana plasmática é capaz de diferenciar dentre os diferentes solutos disponíveis no meio externo promovendo a absorção de uns em detrimento de outros. A base da seletividade não é bem conhecida, mas pode ser explicada em parte pela força do campo elétrico do sítio de ligação/absorção no transportador ou no canal iônico.
- Participação na transferência de energia
A membrana utiliza o ATP para o transporte ativo de íons propriamente dito. Acredita-se que a energia liberada na hidrólise do ATP seja utilizada para formar um “estado de alta energia que ao ser dissipado permite o movimento do íon contra seu gradiente de potencial eletroquímico. A membrana não apresenta apenas atividade ATPásica, isto é, capacidade de hidrolisar o ATP liberando a energia, mas, também, mantém o gradiente de potencial eletroquímico até o momento de executar o movimento do soluto.
Os nutrientes deslocam-se na solução do solo em direção às raízes, quer por movimento na massa de água que está a ser absorvida pelas plantas, processo designado por fluxo de massa, quer por difusão dos locais de maior para os de menor concentração. A velocidade de transporte depende do teor de água no solo, da concentração do nutriente, da velocidade com que o nutriente é absorvido pela planta, da velocidade de difusão do nutriente na água, da temperatura, e de características do solo que influenciam a tortuosidade do percurso e a adsorção dos elementos na matriz.
A absorção de água e nutrientes dependente da formação de um sistema radicular com um comprimento adequado à espécie vegetal. O crescimento das raízes depende de características genéticas, mas é também influenciado por fatores ambientais como a temperatura, o arejamento, o pH, o teor de água e nutrientes, e a compacidade do solo.
O movimento radial dos nutrientes nas raízes pode dar-se nos espaços intercelulares e poros das paredes celulares (apoplasto), ou no interior das células (simplasto). Embora as paredes celulares sejam porosas, permitindo a passagem dos íons, possuem cargas negativas que adsorvem cátions por atração eletrostática.
A componente lipídica das membranas biológicas é impermeável aos íons. Proteínas de transporte vão mediar a travessia das membranas, podendo ser subdivididas em bombas, transportadores e canais iônicos. As bombas vão realizar um transporte ativo, com gasto de energia. As mais bem caracterizadas transportam prótons através das membranas, com consumo de ATP, e geram um gradiente elétrico entre o citosol e o apoplasto que é utilizado para o transporte passivo de cátions por transportadores ou canais iônicos. A entrada de ânions é feita de modo ativo, sendo acompanhados por prótons, processo designado por co-transporte.
A velocidade de absorção dos nutrientes depende da sua abundância à superfície radicular, do tamanho do cátion, da carga do ânion, do pH, da temperatura e do arejamento do solo. O balanço entre cátions e ânions absorvidos vai influenciar o pH da rizosfera, a atividade das bombas de prótons e a concentração de ácidos orgânicos nas plantas. A presença de elevada quantidade de um nutriente no solo pode inibir a absorção de outro, fenômeno designado por antagonismo iônico.
O transporte de água e solutos no xilema deriva do gradiente do potencial de água entre raiz e parte aérea e dá-se geralmente apenas no sentido ascendente. O fluxo resulta sobretudo da transpiração; mas mesmo na sua ausência, a parte aérea continua a receber nutrientes por movimento no xilema devido à pressão de raiz. O transporte no floema depende do carregamento de açúcares e outras substâncias, que são acompanhadas pela entrada de água e fluxo em direção aos locais onde se dá a remoção. As folhas expandidas são o principal local de carregamento e os destinos são as raízes, os ápices vegetativos e os órgãos reprodutores. Em algumas situações, como deficiente absorção de nutrientes, formação de órgãos reprodutores, ou época da queda das folhas, ocorre grande remobilização de nutrientes à custa da degradação de moléculas nas folhas expandidas e transporte dos nutrientes pelo floema para outros locais. Os nutrientes apresentam diferente mobilidade no floema, sendo o cálcio, o manganês e o boro os menos móveis, e os macronutrientes (com exeção do cálcio) os mais móveis.
2.1 Permeabilidade das membranas
A membrana plasmática é uma bicamada semifluídica de lipídios onde estão mergulhados glóbulos de proteína. Para atravessar a membrana, os solutos, podem fazê-lo através da camada lipídica ou através de proteínas (transportador ou canal iônico). Assim, a seletividade ou permeabilidade depende, entre outros fatores do tipo de soluto e da rota metabólica a ser percorrida (via camada lipídica ou protéica), podendo ter, desta forma, componentes passivos e/ou metabólicos.
A membrana plasmática por ser essencialmente apolar, apresenta forte restrição ao movimento de solutos polares (íons por exemplo). A permeabilidade das membranas, depende também em grande parte do peso molecular e da polaridade do soluto em questão. Além destes fatores a permeabilidade da membrana depende da:
- Temperatura – o aumento da temperatura aumenta a vibração das moléculas constituintes da membrana, aumentando a permeabilidade das membranas.
- Energia metabólica – a manutenção da estrutura e da função das membranas importa em gasto de energia metabólica. Assim, qualquer fator que afete a produção de energia (tensão de oxigênio, venenos metabólicos, etc) provoca aumento na permeabilidade das membranas.
- Íons orgânicos – podem apresentar efeitos gerais e específicos. Os cátions monovalentes ao interagirem com o sistema água/macromolécula, promovem a contração das proteínas com o afrouxamento da camada lipídica, aumentando a permeabilidade das membranas. Os cátions divalentes tem efeito exatamente oposto sobre a permeabilidade das membranas. A explicação para este comportamento não está ainda bem clara.
O encontro dos nutrientes com as raízes pode envolver três processos diferentes:
- Difusão: o nutriente entra em contato com a raiz ao passar de uma região de maior concentração para uma de menor concentração próxima da raiz;
- Fluxo de massa: o contato se dá quando o elemento é carregado de um local de maior potencial de água para um de menor potencial de água próximo da raiz;
- Interceptação radicular: o contato se dá quando a raiz cresce e encontra o elemento.
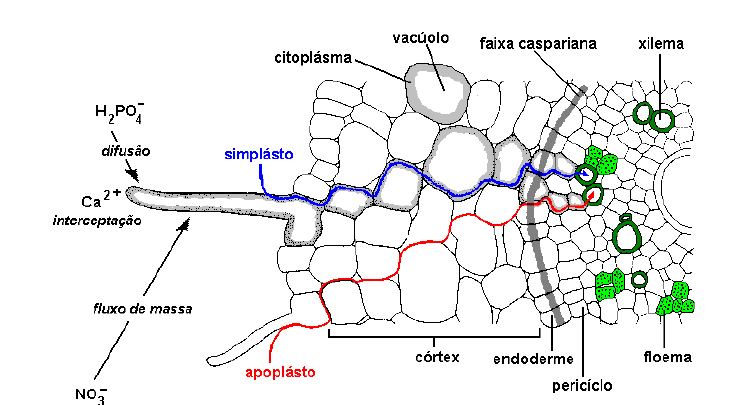 Figura 2: Transporte de nutrientesvia simplasto e apoplasto. O nutriente chega até a raiz (pelo radicular) por difusão, interceptação radicular e fluxo de massa. (Peres, 2002)
Figura 2: Transporte de nutrientesvia simplasto e apoplasto. O nutriente chega até a raiz (pelo radicular) por difusão, interceptação radicular e fluxo de massa. (Peres, 2002)
Os nutrientes muito móveis na solução de solo tendem a chegar até as raízes por fluxo de massa. Um exemplo é o íon nitrato (NO3–), o qual é repelido pelas cargas negativas do solo e por isso tende a se manter solúvel. Por outro lado, o PO43- tende a se ligar a cátions como Fe2+, Fe3+ e Al3+, os quais possuem OH–que é deslocado pelo PO43-. Em conseqüência, o fosfato tende a ser imobilizado pelo solo e tem dificuldade de ser arrastado pelo fluxo de massa. O fosfato chega até as raízes predominantemente por difusão.
A transpiração é importante para os nutrientes que entram em contato com a raiz principalmente por fluxo de massa (nitrogênio, enxofre, magnésio e cálcio). Por outro lado, o tamanho do sistema radicular é muito importante para a absorção de elementos que entram em contato com a raiz por difusão (fósforo e potássio) e interceptação radicular (cálcio).
O crescimento contínuo das raízes é importante para a absorção dos nutrientes. Nem todas as partes das raízes são eficientes na absorção de nutrientes. A zona de maior absorção de íons é a zona pilífera, a qual só está presente em raízes novas como a radícula e as raízes secundárias das dicotiledôneas ou as raízes seminais e nodais das monocotiledôneas. Células dessa zona já se expandiram, mas ainda não possuem crescimento secundário, tendo, portanto uma maior absorção de solutos. Outra questão relevante é que a velocidade de difusão tende a diminuir exponencialmente com o aumento da distância. Desse modo, os elementos próximos das raízes se difundem até elas, mas não são repostos pelos que estão longe, entrando em depleção. Portanto, para uma boa absorção de nutrientes é necessário que o vegetal tenha um crescimento radicular contínuo. A contínua formação de raízes garante raízes novas (absorventes) que alcançam áreas do solo onde o nutriente difundido ainda não entrou em depleção.
O crescimento radicular é controlado geneticamente e depende de uma série de fatores como o estímulo hormonal, a disponibilidade de fotoassimilados e condições do solo como textura, estrutura, umidade, pH e nutrientes. Quanto ao pH, é interessante notar que o crescimento radicular geralmente é favorecido em solos um pouco ácidos, com pH variando entre 5,5 e 6,5. Na década de 70, Drew e colaboradores demonstraram que as raízes tendem a crescer mais na direção onde se encontram os nutrientes. Esse mesmo fenômeno também ocorre com relação à disponibilidade de água. Embora o mecanismo que explique o crescimento preferencial das raízes em direção à água seja relativamente simples, ainda não temos uma explicação fisiológica para o caso dos nutrientes.
Com relação à disponibilidade de água, sabemos que a turgescência é necessária para a expansão celular e como os ápices radiculares perdem turgescência nas porções secas do solo, as raízes crescem preferencialmente nas zonas do solo que permanecem úmidas. Essa é a razão pela qual ao observarmos a direção do crescimento radicular temos a falsa impressão de que a raiz “procura” a água. Uma aplicação prática dos trabalhos de Drew é que, ao fazermos uma adubação superficial ou muito localizada, estaremos estimulando o crescimento das raízes de um modo também concentrado. Esse tipo de crescimento é prejudicial para a planta já que, em condições onde a água ou outros nutrientes possam ser escassos, um sistema radicular profundo e diversificado seria mais vantajoso.
Uma alternativa para facilitar a absorção de nutrientes são as micorrizas. As micorrizas são associações das raízes com fungos que estendem o sistema radicular dos vegetais, sendo muito importante para aumentar o contato com o fósforo. Os nutrientes podem chegar até o xilema das raízes via simplasto ou apoplasto.
Uma vez entrando em contato com as raízes, os nutrientes precisam chegar até o xilema. Após entrar em contato com a raiz, o nutriente pode chegar até o xilema via apoplasto (parede celular e espaços intercelulares) ou simplasto (conjunto de citoplasmas interligados pelos plasmodesmatas). Mesmo para aqueles elementos absorvido inicialmente via apoplasto, para que cheguem até o xilema precisam entrar dentro da célula quando atingem a endoderme. Isto ocorre porque a endoderme apresenta uma barreira ao apoplasto denominada faixa caspariana. No xilema os solutos voltam a cair no apoplasto, já que os elementos de vaso são células mortas.
O processo pelo qual o íon deixa o simplasto e entra no xilema é chamado “carregamento do xilema”. Tratamentos com citocininas (BAP) inibem o carregamento de solutos no xilema sem afetar sua entrada na córtex. Há evidências de que o carregamento do xilema está sob o controle de H+-ATPases e canais de efluxo de íons.
Transporte de Solutos Através de Membranas
As células acumulam alguns nutrientes em concentrações muito superiores à do meio externo, enquanto o nível celular de outros solutos quase sempre é menor que o do meio externo .Portanto, as células devem possuir mecanismos que, especificamente, selecionam aqueles nutrientes que devem ser absorvidos, ou descriminados. Esses mecanismos são operantes em nível de membrana plasmática, que atua como uma barreira efetiva à passagem de íons e moléculas polares não carregadas. Termodinamicamente, o acúmulo de um soluto é um processo não espontâneo e, portanto, a seletividade da absorção e o acúmulo devem envolver uma fonte energética, além de carregadores ou permeases com sítios de ligação específicos para uma determinada espécie iônica, ambos localizados na plasmalema.
A membrana plasmática, bem como outras membranas celulares, controla intenso transporte de solutos para dentro e para fora das células. Os principais sistemas de transporte operando nestas membranas são:
{kind=link}
Nesse tipo de transporte há o gasto de energia (na forma de ATP) e ocorre contra um gradiente de concentração, isto é, as substâncias serão deslocadas de onde estão pouco concentradas para onde sua concentração já é alta. O exemplo clássico de transporte ativo é a BOMBA DE SÓDIO E POTÁSSIO. Os íons de Na+ e K+ são importantes para o funcionamento celular e ocorrem em concentrações específicas dentro e fora das células. O íon Na+ se apresenta em maior concentração no meio extracelular, enquanto o íon K+ se encontra mais concentrado no meio intracelular. Logo o movimento natural desses íons é : o íon Na+ entra na célula por difusão facilitada e o K+ sai da célula pelo mesmo processo. Com isso a tendência é haver um equilíbrio entre as concentrações interna e externa desses dois íons (o que não seria bom para o metabolismo celular), logo a célula gasta energia, na forma de ATP, para fazer o transporte oposto desses íons: colocar o Na+ para fora e colocar o K+ para dentro.
Há dois tipos de transporte ativo:
a) Transporte ativo primário: refere-se ao transporte através de uma membrana contra o seu gradiente de potencial eletroquímico, sempre com gasto direto de energia metabólica (ATP, luz, etc). Este tipo de transporte é intermeado por H+-ATPases das quais são conhecidas três: ATPase “P” (membrana plasmática), ATPase “V” (vacúolo) e ATPase “F” (membrana do tilacóide).
b) Transporte ativo secundário: este tipo de transporte, gasta indiretamente a energia de gradiente eletroquímicos produzidos pelo transporte ativo primário. Pode ser simporte, antiporte ou uniporte.
{kind=link}
É um transporte que ocorre sem gasto de energia externa ao sistema e ocorre a favor do gradiente eletroquímico, ou seja, vai do maior gradiente eletroquímico para o menor gradiente eletroquímico, no caso de um soluto carregado ou a favor do gradiente de concentração, do mais concentrado para o menos concentrado, no caso de um soluto não carregado. Pode ocorrer então a entrada de solutos na célula a partir do meio extracelular ou sair solutos da célula indo do meio intracelular para o meio extracelular.
O transporte passivo ocorre na tendência de equilibrar as forças sobre o soluto. Se for um soluto carregado, existiram forças química e elétrica sobre ele e o fluxo resultante é diferente de zero até o momento em que essas forças se tenham o mesmo módulo e sentidos contrários no soluto carregado. No caso de um soluto não carregado, existe apenas a força química aplicada sobre o soluto e quando a força deixar de existir, pela igualdade das concentrações, o fluxo resultante do íon será nulo. Moléculas hidrofóbicas, alguns gases como o oxigênio e o nitrogênio, por exemplo, conseguem passar facilmente pela membrana e entrar na célula pela membrana. Pequenas moléculas polares não carregadas, como água, glicerol, conseguem entrar na célula mas tem maior dificuldade. Grandes moléculas polares não carregadas, glicose, sacarose, conseguem entrar, mas muito pouco, pela membrana. Íons, como Na+, K+, não conseguem entrar na célula pela membrana. Vale ressaltar que existem os chamados canais iônicos, que são proteínas transmembranas dispostas de maneira a formar um canal, que permitem a entrada dos íons na célula através da membrana.
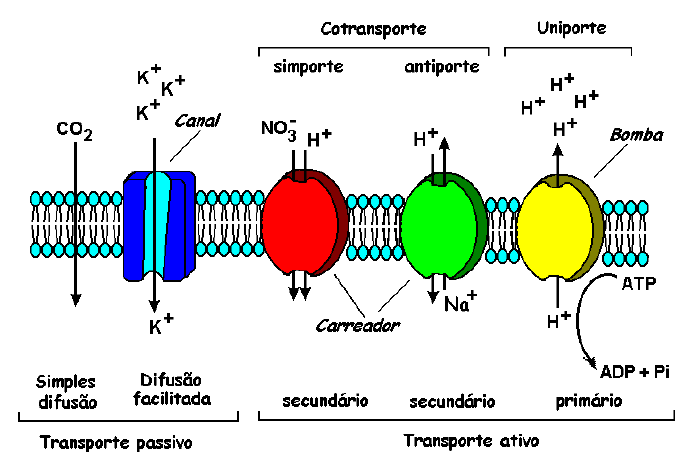
Figura 3: Resumo dos tipos de transporte e da atividade dos carreadores, canais e bombas. Substâncias apolares (CO2) permeiam a membrana livremente. Como a célula vegetal possui um potencial transmembrana negativo, cátions (K+) podem ser transportados por canais, mas os ânions (NO3–) precisam ser transportados por carreadores. O transporte com carreadores utiliza a energia que foi gasta anteriormente pela bomba para criar um gradiente de prótons. Do mesmo modo, a saída de cátions pela bomba para criar um gradiente de prótons. Do mesmo modo, a saída de cátions (Na+) da célula também precisa ser realizada com gasto de energia pelos carreadores. Modificado de Taiz & Zeiger (1998).
Veja: “Bombeamento de Prótons”