
Metabolismo C2
A fotorrespiração diz respeito ao processo em que há absorção de luz associada à liberação de CO2. O termo fotorrespiração significa ainda, que os tecidos fotossintéticos liberam CO2 com maior intensidade na luz do que no escuro, considerando que o processo de respiração (glicólise, ciclo de Krebs e transporte de elétrons) ocorre tanto no período iluminado como no escuro.
A enzima rubisco apresenta-se ativa no tecido fotossintético sob duas formas, uma forma carboxilativa (carboxilase) e uma forma oxigenativa (oxigenase), onde CO2 e O2 competem pelo mesmo sítio da enzima. Diante deste fato, pode-se verificar uma inibição competitiva desses dois gases na fotossíntese, onde o oxigênio se apresenta como inibidor da fotossíntese.
A associação entre a fotossíntese e fotorrespiração foi definitivamente esclarecida com a descrição do processo de oxigenação da RuBP pela Rubisco, concluindo que a relação entre as duas atividades dependia da relação CO2/O2 (condições atmosféricas normais, CO2 = 0,03% e O2 = 21%)ou seja, a inibição da fotossíntese pelo O2 cresce a medida que a concentração de CO2 no ambiente diminui, por esta condição favorecer a atividade oxigenase da rubisco. Em condições atmosféricas normais, a relação entre as duas atividades é de aproximadamente 70:30. Desta competição, decorre uma diminuição da fotossíntese líquida, o que resulta num decréscimo de produtividade nas plantas C3.
O ponto chave do processo está ligado à enzima rubisco presente nos cloroplastos. Ela pode promover a reação da RuBP tanto com o CO2 (função carboxilase) quanto com o O2 (função oxigenase). Quando a concentração de CO2 for baixa e alta de O2, a molécula de O2 não só compete com o CO2, como pode substituí-lo. Como resultado, as duas moléculas de RuBP tornam-se oxigenadas formando duas moléculas de ácido fosfoglicólico (2x2C = 4C) e duas moléculas de 3-PGA (2x3C = 6C) ao invés de quatro, que normalmente seriam formadas na caboxilação (Figura 1).
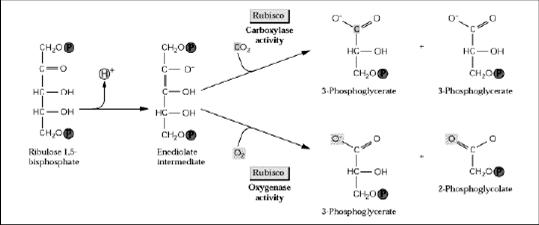
Figura 1. Reação catalisada pela ribulose 1,5 bisfosfato carboxilase/oxigenase (BUCHANAN, 2000).
O processo fotorrespiratório envolve a participação de três organelas, o cloroplasto, peroxissomo e mitocôndria.
O ácido fosfoglicólico (2-fosfoglicolato) por ação de uma fosfoglicolato fosfatase transforma-se em glicolato que se difunde até o peroxissomo onde é oxidado a ácido glioxílico (glioxilato). O glioxilato por ação de uma aminotransferase, produz duas moléculas de glicina que passam para a mitocôndria, onde se convertem em uma molécula de serina (1x3C = 3C) com liberação de CO2. A serina passa para o peroxissomo onde é transaminada a ácido hidroxipirúvico (hidroxipiruvato), que é reduzido a ácido glicérico. O ácido glicérico se difunde até os cloroplastos onde é fosforilado formando o 3-PGA (1x3C). Tanto o 3-PGA quanto aquelas duas moléculas de 2-fosfoglicolato formadas diretamente pela ação da rubisco (no início do ciclo) servirão de substrato para o Ciclo de Calvin. Com o ciclo completo, a estequiometria fica assim estabelecida:
2RuDP + 3O2 + 2FdxH + 3H2O + 2ATP → (3) 3-PGA + CO2 + Pi + 2ADP + 2Fdx
Percebe-se então, que duas das três moléculas de PGA resultam diretamente da ação da RuBP/oxigenase e, a formação de uma terceira molécula de 3-PGA é o resultado do metabolismo de duas moléculas do ácido fosfoglicólico, produzida na mesma reação. Verifica-se assim, que duas moléculas de 2C (ácido fosfoglicólico = 4 átomos de C) são convertidos em uma molécula de 3C (3-PGA = 3 átomos de C) com a liberação de uma molécula de CO2, ou seja, em plantas C3, para cada 2 mol de ácido fosfoglicólico (4C) formado pela ação da atividade oxigenase da Rubisco é perdido um mol de CO2 (1C). Conclui-se daí, que há na fotorrespiração, a recuperação de 75% do carbono que participa em cada “rodada” do ciclo. Os 25% restantes são perdidos para a atmosfera nas plantas C3, como resultado da atividade fotorrespiratória ou são refixados nas plantas C4, como se verá mais adiante.
O metabolismo em plantas C4 inclui também a formação do P-glicolato. Entretanto, nessas plantas não ocorre perda do CO2 pelas seguintes razões:
a) A disposição espacial das células da bainha implica que o CO2 produzido pela fotorrespiração tem que se difundir pelo mesofilo para ganhar o ambiente externo. Todavia, no mesofilo, é fixado novamente pela PEPcase, enzima de alta afinidade por CO2; sendo translocado de volta como ácido dicarboxílico para as células da bainha (C4).
b) O ativo mecanismo de descarboxilação dos ácidos dicarboxílicos nas células da bainha aumenta a eficiência da RuBPcase em detrimento da RuBPoxigenase pelo farto suprimento de CO2, reduzindo-se assim, as perdas de CO2 pela fotorrespiração.
Algumas possíveis funções podem ser pertencentes à fotorrespiração. No contexto da produtividade de biomassa, observa-se que do total de CO2 fotossintético absorvido pela planta, cerca de 18 a 27% em média do carbono é perdido na forma de CO2, sendo este um dos principais fatores de redução na produtividade de biomassa nas plantas C3. Em alguns casos, essa perda pode chegar a 50%. Ao contrário do que possa imaginar, a fotorrespiração apresenta-se como um mecanismo eficiente para as plantas dissiparem energia na forma de calor gerado na etapa fotoquímica, sobretudo sob altas intensidades de radiação, onde os estômatos encontram-se fechados, no sentido de minimizar as perdas de água por transpiração. Esta função, acredita-se ser importante para impedir possíveis danos no aparelho fotossintético.