
A via das pentoses-fosfato ou rota das pentoses é um caminho alternativo do metabolismo da glicose. Esta via não gera ATP, mas exerce algumas funções:
- Produz NADPH para as síntese redutivas, tais como: as biossínteses dos ácidos graxos e dos esteróides,
- Produz ribose para as biossíntese dos nucleotídeos e dos ácidos nucléicos
- Produz eritrose-4-fosfato que combina-se com o fosfoenolpiruvato (PEP) na reação inicial que produz compostos fenólicos vegetais, incluindo aminoácidos aromáticos e precursores de lignina, flavonóides e fitoalexinas
- gera intermediários do ciclo de Krebs.
A via das pentoses-fosfato (desvio da hexose-monofosfato) é um percurso mais complexo do que a glicólise. Trata-se de um processo multicíclico no qual três moléculas de glicose-6-fosfato produzem três moléculas de CO2 e três resíduos de cinco carbonos. Estes resíduos se rearranjam para gerar duas moléculas de glicose-6-fosfato e uma molécula de um intermediário da glicólise, o gliceraldeído-3-fosfato. Desde que duas moléculas de gliceraldeído-3-fosfato possam regenerar a glicose-6-fosfato, a via pode conduzir a completa oxidação da glicose.
3 Glicose-6-P + 6 NADP+ → 3 CO2 + 2 Glicose-6-P + Gliceraldeído-3-P + 6 NADPH + 6 H+
A seqüência de reações da via pode ser dividida em duas fases (Figura 1)
- fase oxidativa, irreversível
A glicose 6-P sofre desidrogenação e descarboxilação formando uma pentose, a ribulose-5-P.
- fase não-oxidativa, reversível
A ribulose-5-P é convertida a glicose-6-P por uma série de reações envolvendo principalmente duas enzimas: a transcetolase e a transaldolase.
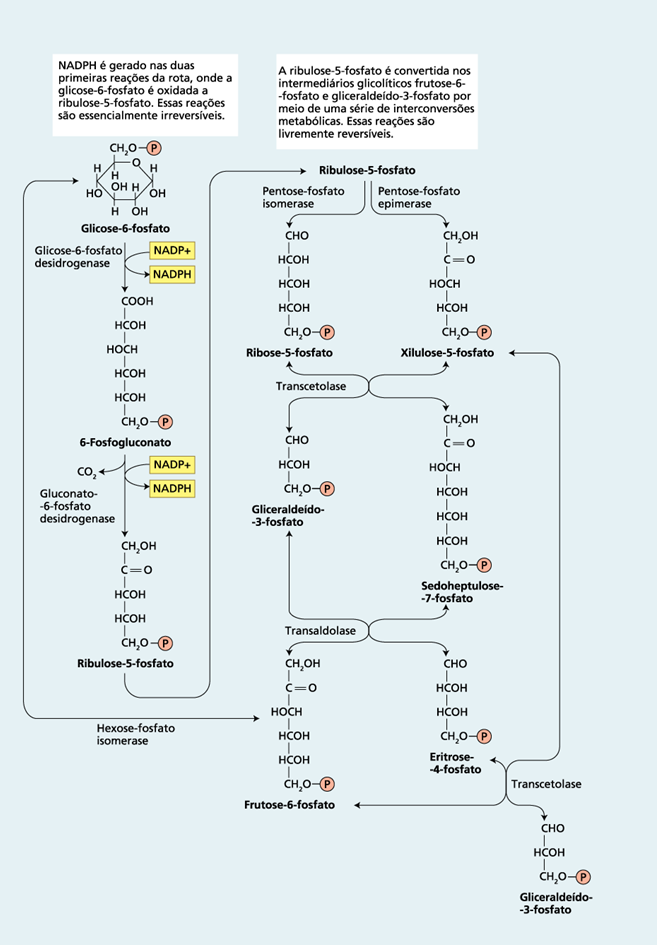
Figura 1: Via das pentoses fosfato
A duas principais vias do catabolismo da glicose são a glicólise e a rota das pentoses, no entanto, essas vias têm pouco em comum. Apesar de alguns metabólitos serem comuns (por exemplo, a glicose-6-P) a via das pentoses é marcadamente diferente da glicólise. Assim, na via das pentoses a oxidação ocorre nas primeiras reações utilizando o NADP ao invés do NAD, e o CO2, que não se forma em toda a via glicolítica, é um produto característico da via das pentoses fosfato, além disso ocorre formação de ribose-fosfato.
O fluxo da glicose 6-P depende da necessidade de NADPH, ribose 5-P e ATP e ocorre basicamente em quatro diferentes caminhos:
- Quando a planta necessita de mais ribose-5-P do que de NADPH, a maior parte da glicose-6-P é convertida em frutose-6-P e gliceraldeído 3-P pela via glicolítica. Então a transaldolase e a transcetolase convertem duas moléculas de frutose-6-P e uma de gliceraldeído-3-P em três moléculas de ribose-5-P pela reversão das reações antes descritas. A estequiometria desse modo de via é:
5 Glicose 6-P + ATP → 6 ribose 5-P + ADP + H+
- Necessidades equilibradas de NADPH e ribose 5-P. A reação predominante nessas condições é a formação de dois NADPH e uma ribose 5-P a partir de glicose 6-P pela via oxidativa da Via Pentose Fosfato. A estequiometria desse modo é:
Glicose 6-P + 2 NADP+ + H2O → ribose 5-P + 2 NADPH + 2 H+ + CO2
- Necessidade muito maior de NADPH do que ribose 5-P, a glicose 6-P é completamente oxidada a CO2. Três grupos de reação estão ativos nessa situação. Primeiro, forma-se dois NADPH e uma ribose 5-P pela via oxidativa da via das pentoses. Então, a ribose 5-P é convertida em frutose 6-P e gliceraldeído 3-P pela transcetolase e transaldolase. Finalmente, a glicose 6-P é re-sintetizada a partir da frutose 6-P e do gliceraldeído 3-P pela via gliconeogênica. As estequiometrias desses três conjuntos de reações são:
6 Glicose 6-P + 12 NADP+ + 6 H2O → 6 ribose 5-P + 12 NADPH + 12 H+ + 6CO2
6 ribose 5-P → 4 Frutose 6-P + 2 gliceraldeído 3-P
4 Frutose 6-P + 2 Gliceraldeído 3-P + H2O → 5 Glicose 6-P + Pi
A soma dessa reações é:
Glicose 6-P + 12 NADP+ + 7 H2O → 6 CO2 + 12 NADPH + 12 H+ + Pi
Assim, o equivalente a uma glicose 6-P pode ser completamente oxidado a CO2 com a concomitante produção de NADPH. A essência dessas reações é que a ribose produzida pela via pentose-fosfato é reciclada para glicose 6-P por transcetolase, transaldolases e por algumas enzimas da via gliconeogênese.
- Necessidade muito maior de NADPH do que de ribose 5-P, a glicose 6-P é convertida a piruvato. A frutose 6-P e o gliceraldeído 3-P derivado da ribose 5-P entram na via glicolítica em vez de reverterem a glicose 6-P. Desse modo geram-se concomitantemente ATP e NADPH, e cinco dos seis carbonos da glicose 6-P emergem em piruvato:
3 Glicose 6-P + 6 NADP+ + 5 NAD+ + 5Pi + 8 ADP → 5 piruvato + 3 CO2 + 6 NADPH + 5 NADH + 8 ATP + 2 H2O + 8 H+
O piruvato formado por essas reações pode ser oxidado para gerar mais ATP, ou pode ser usado como bloco de construção em uma variedade de biossíntese.
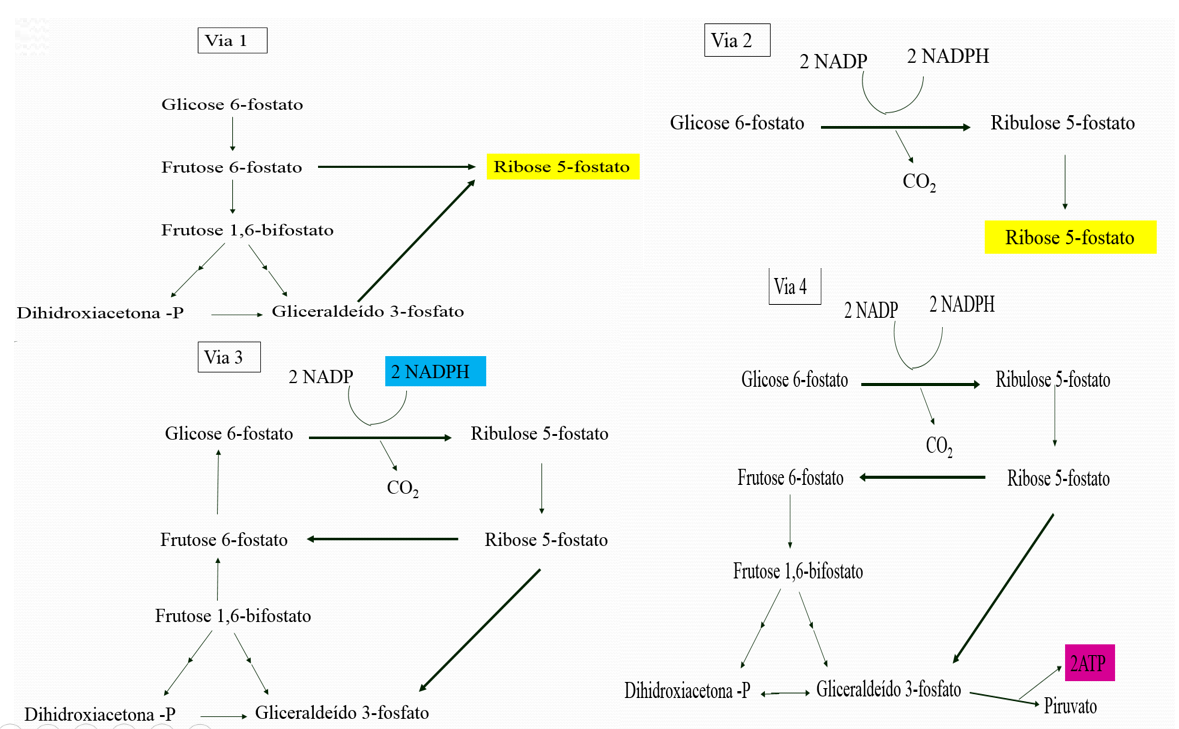 Figura 2: Quatro modo de ação da via pentose fosfato. Os principais produtos são mostrados em cores.
Figura 2: Quatro modo de ação da via pentose fosfato. Os principais produtos são mostrados em cores.
Quando a Via das Pentoses Fosfato assume um papel importante no metabolismo de carboidratos, alternativamente a glicólise?
O pool de triose fosfato/pentose fosfato incluem diversos grupos intermediários de açúcares: ribulose 5-P, ribose 5-P, xilulose 5-P, diidroxiacetona fosfato, gliceraldeído 3-P, sedoheptulose 7-P, eritrose 4-P e frutose 1,6-bisfosfato. A frutose 1,6-bisfosfato é usualmente considerada como parte do pool de hexose fosfato, e também como um substrato para as enzimas transaldolase e transcetolase, funcionando como um componente de ambos pools metabólicos.
A glicólise e a via das pentoses fosfato são ligados por muitas partes intermediárias, não apenas para produzir co-fatores ricos em energia, mas também para gerar esqueleto de carbono requerido para reações biossintéticas. Em resumo, a glicólise, a via das pentoses fosfato e vários caminhos da biossíntese são integrados em plantas pela relação entre o pool de triose/ pentose fosfato.
Na glicólise, após a conversão da frutose 6-P para frutose 1,6-bisP, a reação catalisada pela aldolase da frutose 1,6 bifosfato e isomerase da triose-fosfato são reversíveis. Similarmente, parte da reação da via das pentoses fosfato é também reversível, incluindo o passo catalisado pela epimerase da ribulose 5-P, isomerase da ribose 5-P, transcetolase e transaldolase. Essas reações de reversibilidade serve para manter o pool de triose-P/ pentose-P localizado no citossol e plastídio em equilíbrio.
Existem 3 principais drenos da triose fosfato/ pentose fosfato no metabolismo (Fig. 3).
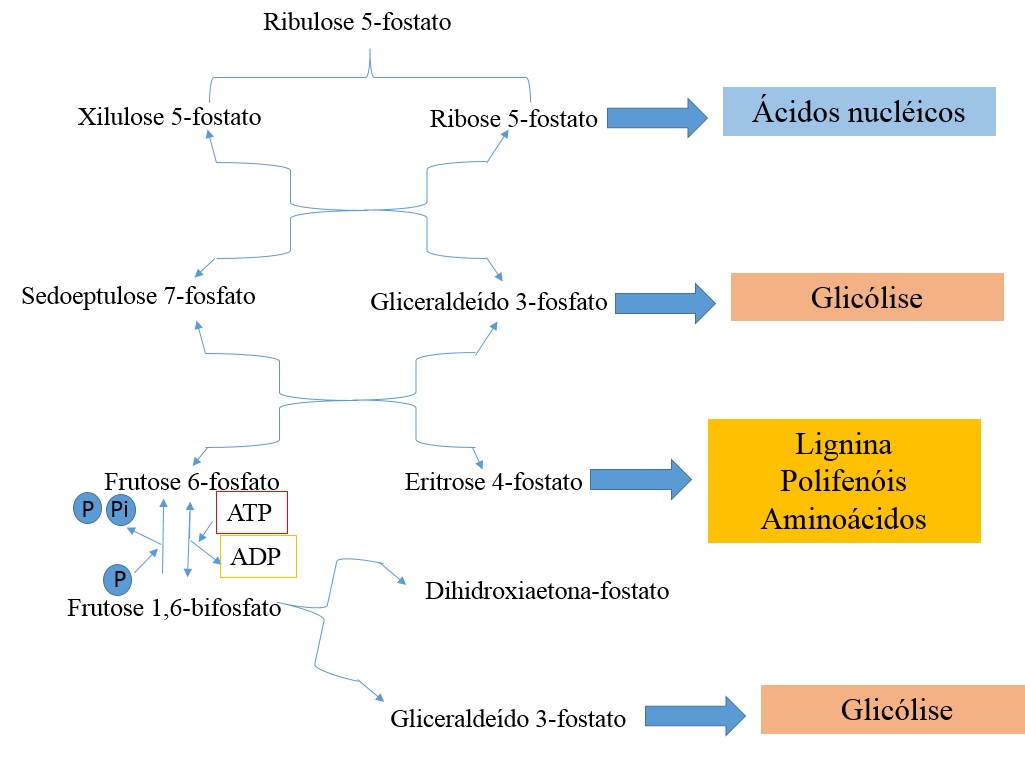
Figura 3. Principais drenos da Via das Pentoses Fosfato
- As trioses fosfato são utilizadas em reações de conservação de energia na glicólise.
- O caminho do chiquimato usa eritrose 4-P para formar lignina, aminoácidos e polifenóis.
- A ribose 5-P é requerido na síntese de ácido nucléico. Quando os metabólitos são drenados para esse pool, as reações reversíveis ajustam para permitir produção dos metobólitos que estão sendo esgotados. Desde de que as reações sejam reversíveis, o fluxo de metabólitos podem ocorrer em ambas direções para formar o produto ou o substrato. As reações oxidativas, na qual não são reversíveis, operam apenas quando o poder de redução é requerido.
As enzimas da parte oxidativa da via das pentoses fosfato são comumente encontradas no citosol e estroma nas células das plantas. No entanto as enzimas da parte não oxidativa tem sido encontradas predominantemente em plastídeos.
Plastídeo do endosperma de feijão, raiz de ervilha, broto de couve-flor e folha de espinafre contém apreciável atividade das enzimas da via das pentoses fosfato. Assim, plastídeos isolados da raiz de ervilha e os cloroplasto de ervilha e espinafre são capazes de reciclar o carbono via pentose fosfato. Estudos da redução do nitrito e síntese do glutamato, estabelece que o fluxo através da via das pentoses fosfato em isolados de raiz de ervilha é depende de reações que requerem o NADPH. Deste modo, o plastídeo na via das pentoses fosfato é considerado como a principal fonte de redução para os processos biossintéticos dessas organelas.
A desidrogenase da glicose-6-fosfato (G6PDH) e desidrogenase do 6-fosfogliconato (6PGDH) são encontrados no citossol em muitas células de plantas. No entanto, recentes estudos sugerem que a repimerase da ribulose 5-P, isomerase da ribose 5-P, transcetolase e transaldolase, na qual catalisam passos na seção não-oxidativa da via das pentoses fosfato, são ausentes no citossol em folhas de espinafre. Isso implica que o citossol em células do mesofilo em espinafre são incapazes da catálise completa da via das pentoses fosfato.
Em tecidos vegetais jovens e meristemáticos a atividade glicolítica é mais intensa e a medida que o tecido vai se tornando maduro a Via Pentose Fosfato se intensifica, para propriciar a deposição de lignina e demais compostos secundários sintetizados com o NADPH.
Outra função da Via das Pentoses Fosfato seria estabelecer a possibilidade de conversão de hexoses, pentoses, tetroses e trioses entre si, com bastante significado econômico nos processos biossintéticos. Portanto a Via das pentoses fosfato além de possibilitar a formação de precursores para biossíntese de metabólicos importantes, fornece energia potencial na forma de NADPH e essa fonte de energá é requerida por exemplo para a assimilação de N, que por sua vez é uma das reações que mais requer esse tipo de energia (Tabela 1). Aumentando o requerimento de energia para a assimilação do NO3-, não é supressa que a redução do nitrato e o metabolismo do carbono são altamente inter-regulados. Devido ao requerimento de carboidratos para a assimilação do nitrato pelas raízes um ajuste na regulação da origem do carbono deve ser importado das brotaçoes. Um ponto pontencial de controle da origem da oxidação do carbono e assimilação do nitrato é a intensa energia da redução da ferredoxina (Fd) requerida pela atividade da NiR e Fd-GOGAT em plastídeos. Em tecidos não-verdes, a FNR (ferredoxina NADP+ oxidoredutase) do plastídeo usa NADPH como substrato para redução da Fd.
Tabela 1. Síntese das principais reações de assimilação de N nas plantas:
| Enzima | Reação | Redutor | Fonte de energia |
| Nitrato Redutase (NR) | NO3– → NO2– | NADH / NADPH | – |
| Nitrito Redutase (NiR) | NO2– → NH4+ | Ferredoxina reduzida | – |
| Glutamina Sintetase (GS) | Glutamato + NH4+ → glutamina | – | ATP |
| Glutamato Sintetase (GOGAT) | a-cetoglutarato + glutamina → 2 glutamatos | Ferredoxina reduzida NADH/NADPH | – |
| Glutamato Desidrogenase (GDH) | a-cetoglutarato + NH4+ → glutamato | NADH | – |
| Asparagina Sintetase (AS) | Gluamina + aspartato → asparagina | – | ATP |
Em cloroplasto, a Fd reduzida é derivada da atividade do FSI durante a exposição à luz. A situação é diferente em tecidos não verdes, onde FNR catalisa o NADPH para redução da Fd para a assimilação do NO2– em aminoácidos. NADPH surge da oxidação da glicose pelas enzimas da via da pentose fosfato (G6PDH e 6PGDH) no plastídeo de raiz. A redução do nitrito é mantida indicando que as pentoses fosfato são recicladas através da via das pentoses fosfato para regenerar glicose 6-P, que então reduz ribulose 5-P com a concomitante produção de NADPH.
Uma possível fonte do NADPH requerido pela redução da Fd em plastídeo de raiz é a oxidação da glicose pela via das pentoses fosfato. Ambos, nitrito e glutamina estimula o fluxo de glicose através da via das pentoses fosfato em plastídeo de raiz de ervilha.
Alguns artigos evidenciam o papel importante que a Via das Pentoses Fosfato no metabolismo de carboidratos:
Artigo 1. Regulação do nitrato pela via da pentose fosfato em plastídeo de raízes de milho: indução da atividade da 6-fosfogliconato desidrogenase (6PGDH)
Nitrate regulation of the oxidative pentose phosphate pathway in maize (Zea mays L.) root plastids: induction of 6-phosphogluconate dehydrogenase activity, protein and transcript levels
Margaret G. Redinbaugh , Wilbur H. Campbell
Plant Science 134 (1998) 129–140
- Efeito do nitrato na atividade da G6PDH e 6PGDH em milho em extrato de raiz e folha:
Para avaliar o efeito externo do NO3– na atividade desidrogenase associado com a via das pentoses fosfato, plântulas de milho crescido hidroponicamente e foram tratadas durante 24 horas com 10 mM de KNO3. A atividade da G6PDH e 6GPDH foi determinada em raízes e folhas tratadas com KNO3 e em plantas controle não tratadas.
Para cada enzima e tecido, a razão significativa da atividade em plantas tratadas com KNO3 em relação as plantas não tratadas foi significativamente maior que 1 (Tabela 2).
Tabela 2. Efeito do tratamento de KNO3 na atividade da G6PDH e 6PDH em milho:
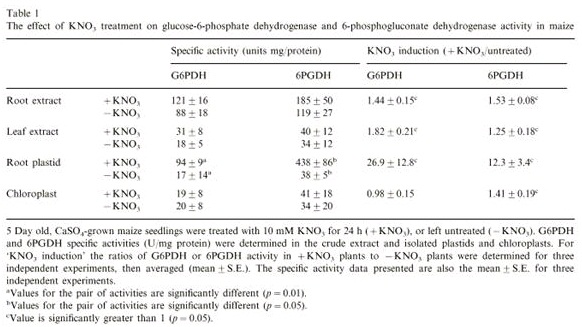
- Efeito do KNO3 na atividade de G6PDH e 6PGDH
Como NO3– foi mostrado previamente aumentando o fluxo do C através da via das pentoses fosfato em plastídeo de raiz, esses autores determinarão o efeito do tratamento com KNO3 na atividade de G6PDH e 6PGDH em isolados de plastídeos de raiz e em cloroplasto de folha. Essas organelas foram separadas do citoplasma baseado na atividade de enzimas marcadas, NR e NiR, respectivamente.
Organelas: plastídeo (raiz) e cloroplasto (folha): NiR
Citossol: NR
A atividade específica das enzimas G6PDH e 6PGDH foi de 27 e 12 vezes maior, respectivamente, em plastídeo isolado de raiz de plantas tratadas com KNO3 em relação ao controle (plantas não tratadas).
A atividade da G6PDH e 6PGDH em isolados de cloroplasto não foi afetado pelo tratamento com KNO3.
Conclusão: O efeito do tratamento com KNO3 na atividade das enzimas na fração do plastídeo de raiz indicou o efeito específico do KNO3 em tecidos e organelas na expressão das enzimas G6PDH e 6PGDH.
Artigo 2. Glicose 6-fosfato desidrogenase como marcador inicial da indução floral em brotações apicais de Espinacia oleracea var Nobel GLUCOSE-6-PHOSPHATE DEHYDROGENASE AS AN EARLY MARKER OF FLORAL INDUCTION IN SHOOT APICES OF Spinacia oleracea VAR. Nobel.
- AUDERSET, P.B. GAHAN a, A.L. DAWSON a and H. GREPPIN
Plant Science Letters, 20 (1980) 109–113 109
O presente trabalho visa determinar como a atividade da via das pentoses fosfato ocorre inicialmente na transferência de plantas para iluminação contínua, e se esse aumento da atividade pode ser anulado pela adicional transferência da planta da luz contínua para o escuro.
Na figura 4 mostra que nenhuma diferença na atividade da G6PDH foi observada entre o controle e as plantas crescidas para 9, 11 ou 13 horas. Após 15 horas de luz contínua alguns ápices mostraram um aumento na taxa da atividade. Após 17 horas todos os ápices mostraram aumento do nível da atividade. A taxa significativa da atividade da G6PDH para todos os ápices iluminados de 17-21 horas foi aproximadamente 2 vezes maior que o controle. Com 15-16 horas de iluminação não foi significativamente afetado pelo retorno para o escuro (8horas).
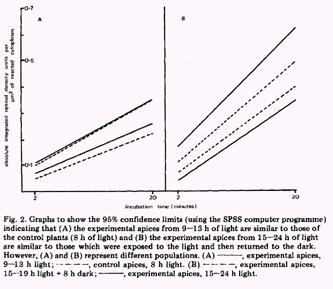
Figura 4
O primeiro aumento significativo da atividade da G6PDH é observado após 15 horas de contínua iluminação com luz branca (para alguns ápices) e com 17 horas todos os ápices mostraram aumento da taxa da atividade da enzima. Esse aumento da atividade da via das pentoses fosfato parece ser o início das trocas do ápice da brotação durante a indução floral. O aumento na atividade da via das pentoses fosfato pode ser relacionado ao aumento frequentemente observado na quantidade de sacarose transportado para o ápice durante o processo de indução.
No entanto, é aceitável que o aumento da atividade da via das pentoses fosfato está ligado a produção de lipídeos através do aumento do nível de NADPH. A via das pentoses fosfato age como uma fonte de precursores de aminoácidos, eles iriam ser necessários nos estágios iniciais do florescimento.
Que relação há entre a Via da Pentoses Fosfato e o ciclo de Calvin?
As plantas verdes contêm em seus cloroplastos uma maquinaria enzimática única que, em um processo chamado fixação de CO2, ou fixação do carbono que catalisa a conversão do CO2 em compostos orgânicos simples (reduzidos). Os vegetais convertem esses produtos simples da fotossíntese em biomoléculas complexas, incluindo açúcares, polissacarídeos e metabólitos derivados dos mesmos. Tomadas em seu conjunto, as reações que promovem a fixação do CO2 integram uma via cíclica na qual os intermediários-chave são constantemente regenerados. Esta via é chamada de Ciclo de Calvin.
Para que o fluxo de CO2 transformado em carboidratos seja contínuo, a ribulose-1,5-bifosfato precisa ser constantemente regenerada. As células vegetais resolveram este problema com uma série de reações que formam uma via cíclica. Por essa via o produto da primeira reação do Ciclo de Calvin (3-fosfoglicerato) passa através de uma série de transformações que eventualmente levam à regeneração da ribulose-1,5-bifosfato.
A regeneração da ribulose-1,5-bisfosfato envolve rearranjos dos esqueletos carbônicos do gliceraldeído e da diidroxiacetona fosfato produzidas nos primeiros dois estágios da fixação do carbono.
Na figura 5, o passo (1) é catalisado pela enzima transcetolase, que contém a tiamina pirosfosfato (TPP) como grupo prostético e requer Mg2+.
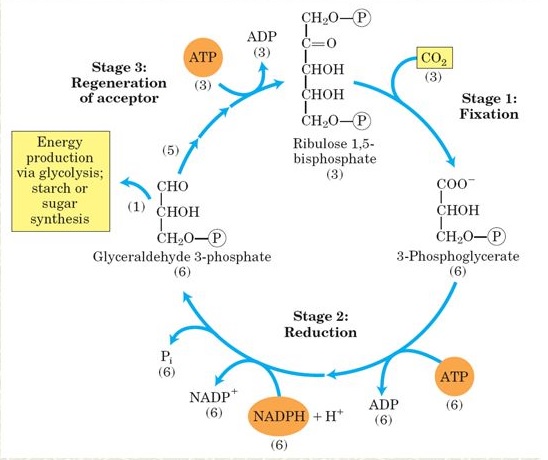
Figura 5. Regeneração da ribulose-1,5-bisfosfato.
A transcetolase catalisa a transferência reversível de um grupo cetol de uma cetose fosfato doadora, frutose-6-fosfato, para uma aldose fosfato receptora, o gliceraldeído-3-fosfato. Os produtos são a xilulose-5-fosfato (uma pentose) e o açúcar de quatro átomos de carbono eritrose-4-fosfato.
O passo (2) é promovido por uma aldolase similar àquela que age na glicólise, ela catalisa a condensação reversível de um aldeído, eritrose-4-fosfato, com a diidroxiacetona na fosfato, produzindo o composto de sete átomos de carbono sedoeptulose-1,7-bifosfato. Depois da remoção do grupo fosfato em C-1 pela sedoeptulose-1,7-fosfato (passo 3), o produto sedoeptulose-7-fosfato é rompido pela transcetolase (passo 4) em uma pentose fosfato (ribose-5-fosfato) e um fragmento de dois átomos de carbono, transportado no TPP. Este fragmento de dois átomos de carbono é condensado com o gliceraldeído-3-fosfato com três átomos de carbono (passo 5) para formar outra molécula de xilulose-5-fosfato em uma reação também catalisada pela transcetolase.
As pentoses fosfato (ribose-5-fosfato e xilulose-5-fosfato) são convertidas em ribulose-5-fosfato (passos 6 e 7), a qual no passo final do ciclo (passo 8) é fosforilada pela ribulose-5-fosfatoquinase e passa a ribulose-1,5-bifosfato.
Em resumo, esta via é a reversão da via das pentoses fosfato e emprega as mesmas enzimas para interconverter hexoses e pentoses fosfato.
A Via da Pentose Fosfato supre o processo fotossintético com NADPH necessário à assimilação do CO2 bem como está intimamente relacionada com a marcha do carbono na fotossíntese.
Durante a fotossíntese, o NADP+ pode ser reduzido pela cadeia de transporte de elétrons, na qual faz desnecessário o passo oxidativo da via das pentoses fosfato. Sobre essas condições, a glicose 6-fosfato desidrogenase localizado no plastídeo é desativada pela fotoredução da tioredoxina.
Em plastídeo de algas verdes Selenastrum, a redução da G6PDH e a fotossíntese são ligadas. Adicionando nitrato na suspensão de células aumenta a demanda do poder redutor, tal que a cadeia de transporte de elétrons da fotossíntese não pode suprir suficientemente o NADPH.
Cloroplasto pode reduzir NADP+ para NADPH usando elétrons da reação de luz da reação de luz na FS ou na reação oxidativa da via das pentoses fosfato. Durante as reações de luz na FS, o fotossistema I reduz a ferredoxina, na qual em voltas, reduz o NADPH e tioredoxina m, uma proteína reguladora. A redução da tioredoxina m transfere H para G6PDH, inibindo a enzima. No entanto, quando outros metabólitos como o nitrato são reduzidos no cloroplasto, a demanda para NADPH aumenta e os elétrons são preferencialmente desviados para a redução do NADP+. Sobre essas condições o pool de tioredoxina torna oxidado e cessa a inibição da G6PDH. Aumentando a atividade dessa enzima ocorre um adicional aumento na produção de NADPH, na qual é necessário para a fixação do CO2, redução SO4– e redução do NO2–.
A glicose 6-P, na qual tem sido importado para dentro de plastídeos não-verde é usado para a síntese de amido e lipídeos ou é alimentado para dentro do plastídeo na via das pentoses fosfato.
A xilulose 5-P é um intermediário do ciclo de Calvin e da via das pentoses fosfato. Ambos ciclos abastece o esqueleto de carbono para outras reações biossintéticas. Por exemplo, a ribose 5-P é usada para a biossíntese de nucleotídeos e a eritrose 4-P é um percurso intermediário da rota do chiquimato principal composto aromático. Sobre algumas condições, mais de 20% do carbono fixado na forma de eritrose 4-P e PEP, e outros percursores da rota do chiquimato são importados dentro do plastídeo via PPT (fosfoenolpiruvato/ fosfato translocador proteína), é direcionado para a síntese de compostos aromáticos.
Tem sido mostrado que a eritrose 4-P pode ser retirada do ciclo de Calvin em grande quantidade sem esvaziar o ciclo. Trabalhos com tabaco demostraram que abastecimento reduzido da Eritrose-4-P, devido a repressão do antisense na atividade transcetolase do plastídio, resulta em uma inibição da fotossíntese e um significativo decréscimo no conteúdo de aminoácidos e fenilpropanoide. Também, mutantes deficientes em PPT mostrou uma forte alteração no metabolismo secundário. Esses dados indicam que o fluxo dentro do metabolismo secundário do plastídio pode ser limitado pelo suprimento de percursores proveniente do metabolismo primário. Sobre condições no qual quantidades significativas de intermediários são removidos da via das pentoses fosfato, a XPT pode abastecer o plastídeo com esqueleto de carbono na forma de xilulose 5-P, gerado no citossol. Isso sugere que a XPT pode representar a posição chave na cooperação entre reações abastecidas de pentose fosfato no citossol e a via das pentoses fosfato em plastídeo. A xilulose 5-P gerado pela sequencia citossólica da via das pentoses fosfato pode ser transportado dentro do plastídeo e alimentado na via da pentose fosfato, na qual produzem NADPH.
Referências Bibliogáficas:
Buchanan, B. B.; Gruissem W.; Jones, R. L. Biochemistry and Molecular Biology of Plants. 1. ed., 2000, p. 696-705.
Taiz, L.; Zeiger, E. Fisiologia vegetal. 5. ed., Artmed, 2012. 918 p.
Artigos: