
Numerosas espécies vivem em zonas áridas e semi-áridas, tendo folhas espessas com uma baixa razão superfície/volume, apresentando taxas reduzidas de transpiração. Estas são plantas suculentas com o metabolismo ácido das crassuláceas- CAM (MARENCO & LOPES, 2009). As plantas CAM representam um grupo de 30 mil espécies vegetais, distribuídas em mais de 25 famílias. Dessas, pelo menos 24 famílias estão no grupo das angiospermas, outras nos grupos das pteridófitas e gimnospermas. Nas angiospermas, as plantas CAM têm sido encontradas nas famílias Cactaceae, Crassulaceae, Orchidaceae, Bromeliaceae, Liliaceae, Euphorbiaceae e outras. Entre as espécies de importância econômica com metabolismo CAM estão o abacaxi, o sisal e as numerosas espécies de cactos e orquídeas. Embora existam plantas CAM aquáticas ou que habitam locais normalmente úmidos, a maioria é encontrada em clima árido. Essas plantas geralmente apresentam folhas espessas e revestidas de uma cutícula densa, o que reduz drasticamente a perda de água, quando os estômatos estão fechados. As células apresentam um vacúolo relativamente volumoso. Nas plantas CAM, os estômatos permanecem fechados durante o dia, para evitar a perda de água, e abertos durante a noite, para permitir a entrada de CO2. Como nas plantas C4, as plantas CAM possuem duas enzimas de carboxilação:
1. a PEPcase, localizada no citosol, que fixa CO2 à noite, formando ácido oxalacético, a partir de HCO3- e PEP;
2. a Rubisco, que atua durante o dia, no ciclo de Calvin, utilizando o CO2 derivado da descarboxilação do malato. O oxaloacetato é reduzido para ácido málico e acumulado no vacúolo à noite. Durante o dia, o malato é descarboxilado para PEP e CO2 ou para piruvato e CO2. O CO2 produzido é fixado pela Rubisco no ciclo de Calvin, durante o dia, como ocorre normalmente nas plantas C3. O PEP utilizado à noite para fixar o CO2 é originado da hidrólise do amido pelas reações da glicólise (MARENCO & LOPES, 2009).
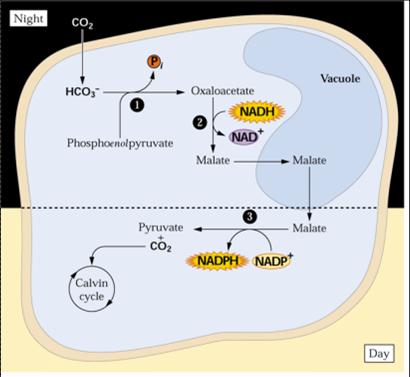
Figura 1. Ciclo das plantas CAM (Buchanan, 2000). 1. PEPcase; 2. Malato desidrogenase- NAD; 3. Enzima málica dependente de NADP.
O comportamento CAM não permite uma grande acúmulo de matéria seca, porém economiza água, podendo então ser considerado como a única via fotossintética que confere adaptação à seca (OSMOND & HOLTUM, 1981). Algumas plantas CAM facultativas podem atingir alta produtividade, quando funcionando como C3, porém quando essas plantas apresentam o comportamento CAM, devido à um estresse ambiental, a acumulação de matéria seca é reduzida.
No metabolismo CAM como as plantas fecham os estômatos durante o dia e abrem durante a noite, elas apresentam pouca transpiração devido ao baixo déficit de pressão de vapor, elas abrem os estômatos, e o CO2 é fixado ao ácido fosfoenolpirúvico pela ação da PEP-case, pois essa reação não depende de ATP ou NADPH. Assim, o carbono é assimilado na forma de ácidos orgânicos, aspártico (ASP) e málico (MAL), acumulados nos vacúolos. Durante o dia, economizam água com os estômatos fechados, pois já assimilaram o CO2, e então descarboxilam estes ácidos orgânicos pelas mesmas vias de descarboxilação C4, fornecendo o CO2 para o ciclo de Benson-Calvin que, este sim, depende da energia luminosa (HALL & RAO, 1994). Essas plantas podem permanecer longos períodos, de 100 a 200 dias sem abrir os estômatos durante o dia, economizando água, porém com baixo acúmulo de matéria seca (OSMOND & HOLTUM, 1981; NOBEL, 1991).
Para a economia de água, além dessa vantagem, as plantas CAM têm uma freqüência estomática (em torno de 2.500 estômatos.cm-2) dez vezes menor que plantas C3 (em torno de 20.000 estômatos.cm-2) (OSMOND et al., 1982).