
Compostos que afetam o transporte de elétrons na fotoquímica
(1) Aceptores de elétrons
Metil viologenio, benzil viologenio, antracnona 2 sulfonato recebem elétrons a partir do PSI em posição anterior a ferredoxina. Ferrocianeto e diclorofenolindofenol (DCPIP), por outro lado, recebem elétrons na região entre o citocromo f e plastocianina.
(2) Doadores de elétrons
Hidroquinona, hidroxilamina e difenilcarbazida doam elétrons entre a água e o citocromo b.
A forma reduzida de DCPIP: N,N,N,N-tetrametil-p-fenilinodiamina (TMPD) doa elétrons no mesmo ponto em que a sua forma oxidada os recebe.
(3) Inibidores do fluxo de elétrons
2 fosfoadenosina difosfato ribose inibe a Ferredoxina-NADP oxidoredutase
(4) Herbicidas
O uso de herbicida para matar plantas não desejadas é difundido na agricultura moderna. Foram desenvolvidas muitas classes diferentes de herbicidas, e eles agem bloqueando aminoácidos, carotenóides, biossíntese de lipídio ou rompendo divisão da célula. O entendimento do modo de ação dos herbicidas foi uma ferramenta importante na pesquisa do metabolismo das planta e facilitou a aplicação deles em práticas agrícolas diferentes.
Alguns herbicidas atuam como inibidores específicos no transporte de elétrons. Vários derivados da uréia, notadamente o Monuron ou CMU (3-p-clorofenil-1,1 dimetilureia), o Diuron ou DCMU [3-(3,4-diclorofenil)-1,1 dimetilureia] e algumas triazinas como antrazina e simazina bloqueiam o transporte de elétrons entre a plastoquinona e o citocromo b.
Paraquat atua recebendo elétrons do PSI na posição da ferredoxina, reduzindo com isto a taxa de produção de NADPH e reduzindo o oxigênio O2 a superóxido (O2-). Superóxido é um radical livre que reage com uma grande gama de moléculas do cloroplasto, conduzindo à rápida perda de atividade do cloroplasto. Moléculas de lipídio da membrana da célula são especialmente sensíveis ao O2-. Como resultado há uma ruptura da membrana do cloroplasto causada por esse radical livre.
Os herbicidas que agem no complexo receptor de quinona competem com a plastoquinona pelo local de ligação QB. Se o herbicida está presente, este desloca a forma oxidada de plastoquinona e ocupa o local específico de ligação do receptor de quinona. O herbicida não pode receber elétrons, assim o elétron não consegue deixar QA, o primeiro receptor de quinona. Assim, a ligação de herbicida efetivamente bloqueia o fluxo de elétron e inibe a fotossíntese. Muitos herbicidas que também agem desta maneira inibem o fluxo de elétron em bactérias fotossintéticas que têm complexos receptores de elétrons do tipo quinona (Figura 1).
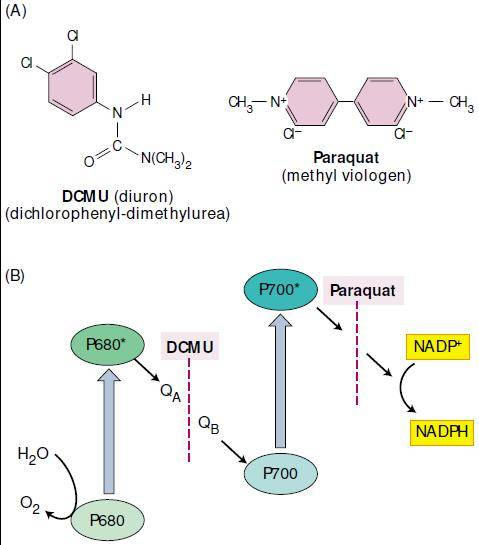
Figura 1. Estruturas químicas (A) e mecanismo de ação (B) dos herbicidas diclorofenil dimetiluréia (DCMU) e metil viologen (paraquat), que bloqueiam o fluxo de elétrons fotossintéticos. O DCMU bloqueia o fluxo nos aceptores quinona do PS-II, por competição com o sítio de ligação da plastoquinona. O paraquat atua recebendo elétrons dos aceptores primários do PS-I (TAIZ & ZEIGER, 2004).
Recentemente, biótipos de herbicidas resistentes de ervas daninhas comuns apareceram em áreas onde um único tipo de herbicida foi continuamente usado durante vários anos. Estes biótipos podem ter ordens de magnitude mais resistentes a certas classes de herbicidas a que plantas não resistentes o são. Em vários casos o fator de resistência foi localizado em uma única substituição de aminoácido na proteína D1. Esta mudança, presumivelmente na região de ligação da quinona (e herbicida) do peptídeo, a diminuição da afinidade de ligação do herbicida, torna este muito menos efetivo.
A possibilidade de uma sintonia da sensibilidade de plantas de cultura aos herbicidas, fazendo mudanças sutis nas proteínas do centro de reação PS-II criou muito interesse na indústria química agrícola. Por biotecnologia, é agora possível tornar uma planta de cultura resistente a um herbicida particular, que pode ser aplicado para controlar plantas indesejáveis que não são resistentes. O sucesso desta modificação dependerá da possibilidade de serem controlados os efeitos colaterais indesejáveis das mutações de herbicidas resistentes e em quão rapidamente as ervas daninhas adquirem resistência por seleção natural ou transferência de gene. Assim, é uma corrida entre a ciência e as mudanças evolutivas.
(5) pH
O gradiente de prótons , o gradiente de pH, através das membranas dos tilacoides é substancial. Quando os cloroplastos são iluminados o pH no compartimento dos tilacoides desce a cerca de pH 5, enquanto no estroma aumenta para pH 8. Uma diferença de 3 unidades no pH é cerca de 1000 vezes na concentração hidrogeniônica. pH = log 1/(H+).