
RESPIRAÇÃO DE MANUTENÇÃO E CRESCIMENTO
A respiração é um processo metabólico complexo e vital nas plantas superiores e produz intermediários de esqueleto de carbono, energia utilizável (ATP) e poder redutor (NAD(P)H) necessários para a maioria dos processos do crescimento e de manutenção. Como resultado, converte uma grande parte do fotossintatos de volta a CO2.
Em relação a respiração de crescimento, podemos dizer que este processo está relacionado com processos biossintéticos, como, incorporação de nova fitomassa e formação de parede e macromoléculas, ou seja, a respiração de crescimento inclui o carbono realmente incorporado – é a conversão de materiais de reservas em novas estruturas: carboidratos estruturais, ligninas, proteínas, ácidos orgânicos.Já a respiração de manutenção fornece a energia para os processos que não resultam em incremento de massa seca, mas sim para manter a funcionalidade celular: manutenção das estruturas de membranas, troca de solutos, substituição de um conjunto de enzimas por outro durante a ontogenia, transporte ativo,reciclagem de macromoléculas (quebra simultânea e re-síntese).
Apesar da importância da respiração dos vegetais para o metabolismo da planta e balanço de carbono na atmosfera e nos animais, alguns de seus pontos chaves são ainda mal compreendidos e a sua quantificação e quantificação bastante difíceis. Uma das metodologias para quantificar e qualificar a respiração dos vegetais, amplamente utilizada e aceita, é através de suas relações com a fotossíntese e o crescimento. Esta é uma área de grande concentração de pesquisa em fisiologia vegetal, principalmente por parte de grupos interessados na quantificação e estimativa da respiração por meio de modelos matemáticos que podem ser aplicados em um bioma ou utilizados para a respiração de uma planta inteira.
Os modelos mais utilizados para a estimativa da respiração são, geralmente, baseados em relações entre respiração e diferentes processos distinguíveis que ela sustenta através de seus produtos. Os dois processos mais claramente distinguíveis (isto é, de maior importância em termos de organização biológica) são o crescimento e a manutenção. Os modelos mais comumente utilizados podem ser agrupados em uma dessas três linhas de trabalho definidas por Amthor (2000) que são mostradas à seguir.
1ª) Produtos da respiração – crescimento e manutenção
A primeira linha de modelagem baseia-se na diferença existente entre adicionar novas moléculas á quantia total de proteínas, lipídeos, celulose, minerais, entre outros (crescimento) e turnover de macromoléculas, transporte de íons intramembranas, entre outros (manutenção). Nesses modelos, há um reconhecimento de que o crescimento e a manutenção são fundamentalmente diferentes. Além disso, classifica todos os processos metabólicos em duas classes: aqueles que são sustentados pela respiração de manutenção e aqueles que são sustentados pela respiração de crescimento, embora o crescimento e a manutenção compartilhem algumas reações bioquímicas.
2ª) Produtos da respiração – crescimento, manutenção e perdas
A segunda linha de modelagem considera as duas variáveis anteriormente observadas (crescimento e manutenção), e adiciona uma terceira variável relacionada às perdas que ocorrem na respiração, ou seja, processos respiratórios que não trazem benefícios para a planta em termos de crescimento ou manutenção. A respiração desperdiçada produz CO2 e/ou calor, mas não contribui diretamente ao crescimento ou a manutenção, com energia e/ou poder redutor. Rotas alternativas de produção de ATP e hidrólise de macromoléculas são consideradas aqui como perdas da respiração, como exemplo temos a atividade da oxidase alternativa mitocondrial.
3ª) Produtos da respiração à relações específicas com cada processo bioquímico
A última linha para a determinação de modelos possui algoritmos que reconhecem as relações individuais que existem entre a respiração e cada processo bioquímico distinguível que ela sustenta, incluindo as perdas. As equações geradas por esses modelos consideram coeficientes de manutenção e de crescimento, que envolvem também as perdas, para cada processo considerado como crescimento ou manutenção, englobando os custos e as taxas de ocorrência de cada processo, podendo assim, explicar processos metabólicos individualmente
Contudo, o mais importante desses modelos é que eles relacionam a respiração (definida em seu sentido mais amplo de troca de CO2 e O2) com a bioquímica básica e fisiologia, dando oportunidades para fazer isto tanto por cálculos quanto empiricamente, embora muitos aspectos da respiração e processos que ela sustenta permaneçam incertos. Este modo de tratar a respiração e os processos relacionados de uma forma individual permite descrever (ao invés de somente explicar) as taxas de respiração observadas em relação aos processos em questão. Assim, por esses modelos pode-se responder questões como: Quanto crescimento poderia ocorrer de uma unidade de fotossíntese? O enfoque desse manuscrito será dado a modelos dessa última linha de trabalho.
Por muitas razões a fotossíntese é uma consideração importante para todos as três linhas de modelagem, porque, basicamente, supre substratos de carbono usados na respiração de crescimento e manutenção. Contudo, a relação entre a fotossíntese e a respiração pode ainda ser mais direta. Por exemplo, a fotossíntese poderia suprir ATP, NAD(P)H e esqueletos de carbono a processos normalmente sustentados pela respiração, destacando alguma respiração nas células fotossintetizantes.
Modelos para estimar a Respiração
Apesar das várias críticas aos modelos matemáticos para quantificar e caracterizar a respiração, deve-se a eles a associação da respiração com o crescimento e produção das plantas . O fato da respiração estar ligada ao crescimento e produção da planta causou a verificação de que fatores que estimulam o crescimento aumentam simultaneamente a respiração. Isto fez com que a respiração passasse de um processo, até então, tido como maléfico à produção e crescimento, pois era responsável direto pela perda de matéria seca na forma de CO2, a um processo essencial para o crescimento e manutenção das células vegetais, pois fazia mais do que apenas liberar CO2 e calor, mas que era essencial para a produção de energia e o abastecimento das rotas de biossíntese.
Os primeiros algoritmos para determinar as variáveis relacionadas à respiração dos vegetais possuíam a forma mostrada na Figura 1. Dentre as várias equações geradas a partir desses algoritmos, a mais simples e amplamente utilizada em todos os modelos, é descrita como :
R= RG + RM= grG + mrW
Onde: R= taxa respiratória (por exemplo, mol de CO2 s-1), Rg = taxa de respiração de crescimento (mol CO2 s-1), RM = taxa de respiração de manutenção (mol CO2 s-1), W= biomassa viva (g biomassa seca), gR = coeficiente de respiração de manutenção (CO2 liberado devido ao crescimento por unidade de nova biomassa produzida ex. mol de CO2 g nova biomassa s-1), G = taxa de crescimento (g nova biomassa s-1), mr = coeficiente de respiração de manutenção (CO2 liberado devido à manutenção por unidade de biomassa existente, ex. mol de CO2 g de biomassa viva-1 s-1).
Alguns comentários sobre a equação são pertinentes. O gR é uma relação que representa o subproduto de CO2 do crescimento enquanto mR é uma taxa associada com atividades de manutenção. O gR e o mR podem ser estimados empiricamente, bastando para isso medir simultaneamente R e as outras variareis ou, podem ser calculados a partir de dados dos processos metabólicos como será mostrado mais adiante (Cannel & Thornley, 2000; Gifford, 2001).
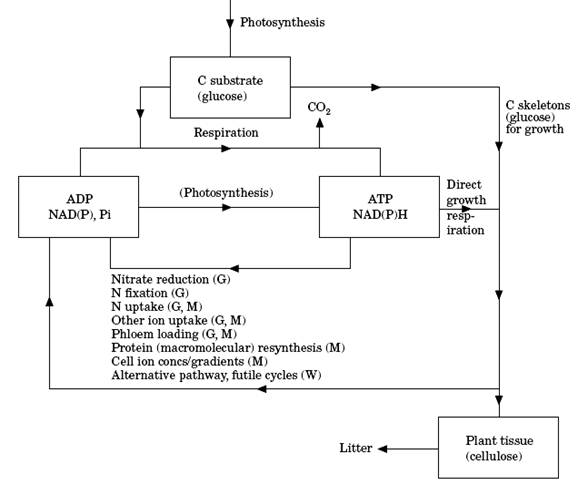
Figura 1. Esquema simplificado do fluxo de carbono destinado ao crescimento (G), à manutenção (M) e pardas (W). Fonte: Cannell & Thornley (2000).
A questão das prioridades para o uso de fotoassimilados por meio dessa equação fundamental é às vezes levantado por alguns pesquisadores. Por exemplo, para um determinado processo existe uma taxa fixa de respiração de manutenção exigida ou necessária, então como o crescimento é sustentado pelo substrato que resta da manutenção?
De modo dinâmico, as plantas equilibram o uso de substrato fotossintetizado entre manutenção e crescimento, dependendo das condições ambientais, estado fisiológico e estado de desenvolvimento. Estas implicações de prioridades do uso de substrato para manutenção e crescimento, dentro do contexto de modelos matemáticos, foram recentemente avaliados por Thornley e Cannel (2000). Em seu trabalho apontam que a equação mostrada acima, como a maioria das outras equações de respiração, não específica prioridades, ela apenas mostra que o RG e o RM constituem a respiração nas plantas em crescimento.
Por outro lado, alguma fração da taxa de manutenção é continuamente necessária em células vivas e, portanto, a manutenção desempenha uma certa prioridade mínima para uso de substrato. Mas, como os coeficientes mR e gR são variáveis em relação ao tempo e condições ambientais, as prioridades aparentes podem igualmente variar.
É difícil separar perdas de manutenção. Thornley e Cannel (2000) notaram que as perdas da respiração poderiam aumentar o gR e o mR aparentes, dependendo de sua natureza bioquímica. Se cálculos determinam o que o gR e mR devem ser, estes valores poderiam ser comparados as medições destes coeficientes (baseado na equação mostrada) para estimar o grau de perda. Dessa maneira, até o ponto que alguma respiração é perdida, a equação fundamental mostrada acima pode ser considerada incompletos.
Um ponto importante para a determinação dos coeficientes de crescimento e de manutenção e sua relação com perdas é que; a produtividade máxima de uma unidade de fotoassimilado seria obtida se o ATP e o NAD(P)H produzido pela respiração dessa unidade fossem usados apenas em reações que contribuíssem diretamente para o crescimento e manutenção. Essa produtividade pode ser medida pela relação de produção de ATP (a partir de ADP e Pi) pela liberação de CO2 na respiração, o que dá idéia da eficiência respiratória relacionada à produtividade de energia por unidade de fotossíntese. A relação de ATP produzido por CO2 liberado na respiração é dada por:
Y ATP,C [mol de ATP mol CO2-1]
Onde: YATP,C é a produção de ATP por CO2 liberado em um processo bioquímico caracterizado como manutenção ou crescimento. A importância do índice YATP,C para a determinação dos índices mR e gR de uma rota ou processo qualquer é poder determiná-lo por meio de cálculos ou pelo simples conhecimento das rotas metabólicas em questão (Amthor, 2000).
O valor máximo de YATP,C de um processo que consome bastante energia pode ser um pouco menor do que 5 (mol de ATP mol CO2-1) ao passo que em alguns estudos mais antigos foram observados valores máximos de YATP,C da ordem de 6 a 6,3 (mol de ATP mol CO2-1).
Respiração de manutenção
A definição de manutenção de Penning de Vries et al. (1983) permanece útil até os dias de hoje. Segundo estes, além de outros autores a manutenção de uma planta inclui processos que mantém as estruturas celulares e gradientes intracelulares de íons e metabólitos, juntamente com ajuste fenotípico ás mudanças ambientais. A substituição de um conjunto de enzimas por outro durante a ontogenia pode também ser considerada como manutenção. Processos de manutenção dominantes em uma célula são: reciclagem macromolecular (turnover) e transporte ativo (Penning de Vries et al., 1983). A Tabela 1, mostra a classificação de alguns processos em. crescimento, manutenção e perdas; note que alguns processos possuem mais de uma classificação, dependendo da linha de trabalho de modelagem.
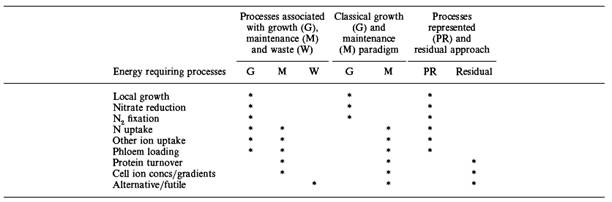
Tabela 1. Diferentes classificações e definição de processos que requerem energia nos vegetais Fonte: Thornley & Cannell (2000)
A respiração de manutenção é a liberação de CO2 resultante de atividades dos processos de manutenção. Os processos de manutenção podem consumir principalmente ATP ao invés de esqueletos de C ou NAD(P)H . Voltando à equação básica [ R = Rm + Rg], a taxa de respiração de manutenção RM pode ser calculada à partir de taxas de processos fundamentais se os custos metabólicos e estequiometrias de liberação de CO2 desses processo forem conhecidos .
As questões se encerram então em verificar quais são as taxas de processos de manutenção e quais são os custos metabólicos em unidades de CO2, ou glicose, de tais processos. Resolver tais questões é simplesmente uma abordagem mecânica da avaliação do coeficiente de respiração de manutenção (mR). Com base nas taxas e nos custos dos processos de manutenção, o coeficiente mR pode ser decomposto para explicar diferentes processo de manutenção, conforme a equação abaixo:
RM=∑processos,X mRX = ∑processos,X cX aX
onde, X é um processo de manutenção qualquer, mRx é o coeficiente de respiração de manutenção para o processo X; cX é o custo do processo X (em CO2 por unidade de atividade de x) e aX é a taxa do processo x por unidade de biomassa (atividade especifica).
Como exemplo dessa abordagem mecânica para o cálculo de mR, dois processos (turnover de proteínas e transporte de íons intracelular) serão abordados e exemplificados, com base no trabalho de Amthor (2000).
1 – Turnover de componentes celulares
A degradação de proteínas é, de modo geral, catalisada por proteases sob regulação metabólica. O turnover protéico permite às células alterarem a sua constituição metabólica em resposta a ontogenia e/ou a mudanças ambientais, o que facilita a remoção ou a substituição de proteínas anormais ou danificadas e também substitui complexos enzimáticos. Sem o processo de turnover, as exigências protéicas seriam elevadas devido a constante síntese protéica em uma faixa de condições ambientais e todos os estágios de desenvolvimento. Respostas rápidas a mudança ambiental ou estresses podem exigir um nível bastante elevado e turnover rápido, embora sejam poucas as evidencias de que a taxa de turnover de proteínas deva ser rápida .
O ATP exigido por aminoácido pra o turnover de proteína é estimado na Tabela 2. A conversão para custo de turnover protéico (cpt) (em CO2 / aminoácido) depende da relação de liberação de CO2 por ATP formado ou seja, 1/YATP,C. O valor mínimo (mais eficiente) dessa relação é cerca de 0,2 CO2/ATP. Isto dá um cpt 0,9 – 1,6 CO2 / aminoácido, para o caso de reciclagem completa da proteína (quebra e síntese), com um custo de ATP de 4,7 –7,9 por aminoácido. Note que cpt inclui custo de turnover de RNA, que é bastante reduzido em relação aos outros custos. O turnover de outros RNA’s é provavelmente uma fração ainda menor de cpt e podem ser, para efeito de cálculos, desconsiderados .
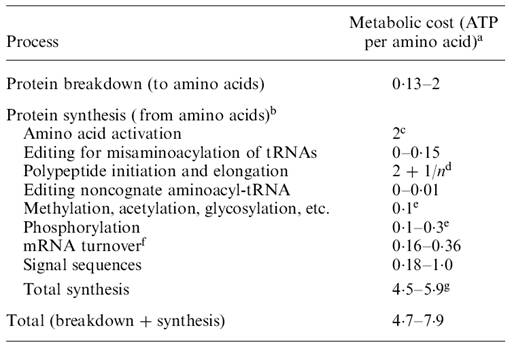
Tabela 2. Estimativa de custos específicos para o turnover protéico.
As taxas de turnover protéico são bastante variáveis entre espécies, órgãos e ambientes bem como sazonalmente. Existem relatos indicando que, diariamente, cerca de 6,5 a 21% da proteína total de uma célula é degradada e sintetizada de novo. Membranas celulares, incluindo suas proteínas, também possuem turnover. A plasmalema de algumas células podem sofrer o processo de turnover a cada poucas horas, embora nenhum custo metabólico deste processo tenha sido estimado até hoje.
Manutenção do gradiente intracelular
O transporte ativo de íons para regular pH ou potencial osmótico das células é parte da manutenção, a compartimentalização dos íons é parte do crescimento. Para avaliar o custo de transporte ativo de íons (cíon = CO2/íon) a liberação de CO2 deve estar relacionada estequiometricamente á fonte de energia de transporte. Essa fonte pode ser ATP, mas pode também ser fosfato inorgânico (Pi) nos tonoplastos e talvez NAD(P)H nas plasmalemas.
Usando ATP e considerando-se a relação H+ : ATP = 1:1 e íon : H+ = 1:1, cíon é igual a 1/YATP, C. Diferentes valores para cíon podem surgir caso a fonte de energia seja Pi ou NAD(P)H com a mesma relação íon : H+. Baseado nos dados de fluxo de íons de condições de laboratório, Penning de Vries et al. (1983) observaram uma ordem de magnitude de transporte ativo de íons de 2 mol de íon kg-1 biomassa dia–1.
Com cíon = 0,2 CO2/ion (YATP,C máximo), o coeficiente de manutenção de gradiente iônico intracelular mRíon é igual a = cíon x aíon = 400 mmol de CO2 kg-1 biomassa d-1.
Medindo o mR
Além de calcular o mR (ou os seus componentes) a partir dos custos e taxas de processos básicos, ele pode ser estimado medindo a taxa de respiração R. Por exemplo as equações demonstradas podem ser solucionadas experimentalmente, porém, para isso, é necessário cuidado quando se usa resultados individuais, porque vários fatores podem comprometer a precisão das medidas de respiração.
Penning de Vries (1975), entre outros, propôs medir a razão R/W no escuro como um método seguro para estimar mR. Quando uma planta é colocada no escuro, há esgotamento de suas reservas e o crescimento para. Neste ponto, o efluxo de CO2 seria totalmente devido a manutenção. Devido a sua simplicidade este método foi usado com freqüência, mas, pode ser um método pouco confiável. Isso porque, durante períodos prolongados de escuro, a funcionalidade fisiológica pode diminuir, invalidando a suposição de que a respiração esteja refletindo custos de manutenção normais.
Outro método de avaliar o MR é medir a razão R/W em tecidos ou órgãos maduros. A suposição é que órgãos maduros não crescem, assim RG = 0 e RM = R. Uma complicação é que mesmo em órgãos maduros os processos de não manutenção podem ocorrer. Para as folhas (órgão favorito de estudo) a dificuldade mais clara diz respeito a respiração que sustenta a translocação de fotoassimilados e a senescência. Este método é não obstante popular para estimar o mR foliar. A sua atração é que não envolve os tratamentos especiais ou as condições experimentais, simplesmente medições de respiração de órgãos intactos .
Duas generalizações sobre o mR são bastante relevantes: a primeira é a de que está associado a temperatura e outra é que reponde positivamente ao teor de nitrogênio (N) da planta. Alem das associações com padrões de temperatura e com o conteúdo de N, mR pode estar positivamente relacionado com a taxa metabólica total, avaliada em termos da assimilação de CO2 líquida. Esta propriedade do mR foi incluída em modelos como um componente separado do RM junto com turnover protéico e componentes de manutenção de gradientes de íons.
Respiração de Crescimento
O crescimento, em termos de modelagem da respiração vegetal foi definido de muitos modos. A definição mais usual é a de que crescimento é a conversão de materiais de reservas (carboidratos não estruturais, lipídeos, entre outros) em nova estrutura, isto é: carboidratos estruturais, ligninas, proteínas, lipídeos, ácidos orgânicos etc; ao invés de simplesmente mudança na massa seca total.
Em principio a respiração de crescimento pode ser fornecida, diretamente, ao se calcular o CO2 liberado e o substrato consumido durante o crescimento, simplesmente somando-se todas as reações bioquímicas de crescimento e equilibrando as exigências de ATP e NAD(P)H liquidas com a quantia de ATP e NAD(P)H produzidos por substrato respirado. Este método de analise de rotas metabólicas para calcular o gR exige conhecimento dos substratos (açucares e amidas especificos usados no crescimento), rotas de biossíntese e composição de biomassa produzida no crescimento .
O gR é independente dos fatores ambientais, como a temperatura, até o nível que a quebra dos substratos, rotas de biossíntese e composição de biomassa são igualmente independentes desses fatores. Além disso, o método de cálculo direto do gR não prediz a taxa de crescimento G, sendo então necessário o conhecimento separado de G para calcular a taxa de respiração de crescimento RG (= gR x G).
O aspecto chave desse método é o calculo de gR a partir da bioquímica básica. Uma limitação é a dificuldade de obter dados de composição de biomassa completos e exatos. Além do mais, o conhecimento das rotas é as vezes incompleto, especialmente, para compostos secundários. Por exemplo, Penning de Vries (1975) estimou a rota de síntese de lignina por que dispunha de descrições incompletas. De maneira semelhante, a síntese de hemicelulose e de alguns componentes da biomassa estavam ou foram muito simplificados nas suas análises. O conhecimento das rotas biossintéticas progrediu desde então e o método tem sido aplicado a uma diversidade maior de componentes e biomassa, embora permaneçam pontos obscuros sobre algumas rotas.
Penning de Vries (1975) simplificou este método de calcular os custos de crescimento por rotas metabólicas categorizando os compostos em 5 grupos: compostos nitrogenados (principalmente proteínas e amino ácidos), carboidratos (principalmente estruturais), lipídeos, lignina e ácidos orgânicos. Diferenças em custos de biossíntese entre compostos dentro de grupos foram consideradas pequenas o bastante para serem desconsideradas, porém diferenças entre os grupos são grandes. Esta simplificação permitiu a aplicação do método para composição aproximada de biomassa (isto é fração de biomassa composta de carboidratos, proteínas, lipídeos, ligninas e ácidos orgânicos) ao invés de exigir dados de composição mais detalhados e difíceis de obter.
Além de estabelecer os 5 principais grupos que consomem energia para o crescimento, o mesmo autor identificou 5 sub processos do crescimento que consomem energia e/ou esqueletos de carbono: redução do NO-3 e SO2-4; absorção ativa de minerais e substratos orgânicos em células em crescimento, síntese de monômero a partir destes substratos, polimerização, manutenção de ferramentas, absorção ativa de minerais pelas raízes e o carregamento do floema em órgãos fontes que suportam o crescimento e uso de energia.
A redução química de qualquer NO-3 e SO2-4 absorvido do solo exige agentes redutores. Estes são formados na respiração (e/ou fotossíntese em células fotosinteticamente ativas) A absorção ativa de minerais e substratos em células em crescimento exige presumivelmente ATP e, esse ATP é derivado principalmente da respiração.
A polimerização de alguns monômeros exige energia na forma de ATP ou agentes redutores. Essas podem ser derivadas da respiração (e/ou fotossíntese) O esboço de ATP dado na Tabela 2 se aplica ao crescimento bem como a manutenção. A manutenção da ferramenta é o turnover do RNA e enzimas que catalisam o crescimento. É distinguida da manutenção destacada acima que foi chamada de manutenção de estrutura porque é dependente da taxa de crescimento. Os seus custos, que são provavelmente uma pequena fração dos custos de crescimento total são calculados como na Tabela 2.
A absorção ativa de minerais pelas raízes exige energia (ATP) que é produzido pela respiração. O carregamento do floema em órgãos fontes também exige energia na forma de ATP, que é produzido pela respiração (e/ou fotossíntese durante o dia). A síntese de monômero é vital ao crescimento e constitui o uso principal de substratos durante o crescimento. Por esse processo é possível explicar a conservação de carbono dentro da nova biomassa formada.
Na respiração de uma planta toda, a equação geral da respiração relaciona gR com a quantidade de CO2 liberado por unidade de crescimento. Para plantas inteiras, todos os sete sub processo de crescimento estão ou são incluídos no gR. Porém, para órgãos individuais, o gR inclui apenas transporte ativo, síntese de monômero a partir de substâncias importadas tais como sacarose e amidas, polimerização e manutenção de ferramentas. É, portanto útil considerar um gR que descreve a respiração de crescimento dentro de órgãos em crescimento. Processos relacionados a crescimento excluídos do gR dos órgãos individualmente, tais como assimilação de NO-3, absorção de íons do solo e carregamento do floema podem talvez ser melhor tratados como componentes respiratórios.
A relação respiração/fotossíntese
A abordagem da relação entre a respiração e a fotossíntese é remissiva à relação existente entre massa e energia. Em termos demassa (seca) de produtos sintetizados por unidade de massa (seca) de substrato usado, lipídeos são caros ao passo que carboidratos estruturais são baratos, como pode ser observado na Tabela 3. Mas, em termos de energia fixada em produtos por energia de substrato, há pouca diferença entre os compostos, especialmente quando se compara quantidades semelhantes de produtos vegetais (Pereira, 1989). Além disso, o teor de carbono da biomassa é positivamente relacionado ao conteúdo energético (através do estado de redução) em todas as macromoléculas .
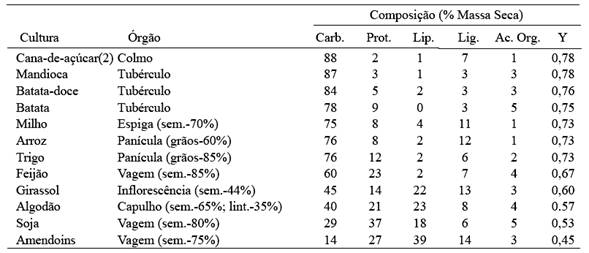
Tabela 3. Composição média e eficiência de conversão (Y) de algumas culturas Fonte: Pereira (1989).
Um modelo simples e muito eficiente que permite quantificar a utilização e distribuição de substrato da fotossíntese nos processos de crescimento é bastante utilizado. A quantidade total de substrato (ΔS) produzido pela fotossíntese bruta (Fb), num intervalo de tempo (Δt), é dada por:
ΔS = Fb x Δt
A fotossíntese bruta, no entanto, pode ser descrita como a diferença entre o montante de carbono que é fixado e o montante desse elemento que é liberado pelos processos respiratórios. Teoricamente, e como já largamente discutido, o substrato reduzido e fixado pela fotossíntese (ΔS) pode ser utilizado em dois processos respiratórios distintos: o crescimento (ΔSc) e a manutenção (ΔSm), equacionados como:
ΔS =ΔSc + ΔSm.
Ainda, como anteriormente visto, a manutenção é obrigatoriamente exigida em todas as células vivas da planta e, por isso, o substrato destinado para a manutenção (ΔSm) é totalmente respirado e, por depender do montante de células vivas, essa variável é dependente da biomassa existente (W).
ΔSm = M * W * Δt
Onde: M é o coeficiente de respiração de manutenção.
Porém, essa não é a única forma de respiração de manutenção, pois parte do substrato destinado ao crescimento (ΔSc) também é respirado para suprir os custo de síntese de compostos para o crescimento (Cc), liberando energia para converter o restante do substrato destinado ao crescimento em nova biomassa (Ce), assim:
Sc = Cc + Ce
A eficiência (Y) do processo de conversão de substrato fotoassimilado é dada pela relação entre o incremento da biomassa (Ce) e quantidade de substrato disponível para crescimento (ΔSc), isto é:
Y = Ce / ΔSc = Ce / (Ce + Cc)
Assim, o teor de carbono da biomassa está inversamente relacionado com o Y.
O Y pode ser expresso com base em massa (carbono ou glicose) ou em energia retida nas ligações químicas (ATP). Em alguns contextos ecológicos, um Y baseado em energia pode ser mais importante do que o Y baseado em massa, porém, o teor de energia é também uma medida imperfeita do rendimento útil dos processos de crescimento. Por exemplo, os grupos aminas das proteína não podem ser oxidados pelos animais, assim embora um pouco da energia no substrato seja retida neles, essa energia não está disponível aos animais. Isso também ocorre com a celulose, que tem Y com base em massa e em energia altos, mas não podem ser usados como fonte de carbono ou energia por muitos animais.
A eficiência de conversão (Y) pode também explicar o porque de não ser observada uma relação direta entre fotossíntese e produção vegetal. Em geral, o aumento na produtividade resulta na seleção de plantas menos competitivas, mas com maior capacidadede alocação de produtos fotossintetizados aos grãos. Portanto, uma mesma taxa de fotossíntese acarreta taxa de crescimento inversamente proporcional ao tamanho da planta, em função da respiração de manutenção. Isso explica, em parte, a maior produtividade em variedades de porte reduzido, que, em tese, possuem uma menor respiração de manutenção. Por esse ângulo, o crescimento pode ser estimado simplesmente medindo-se a relação entre a respiração e a fotossíntese das plantas.
Uma questão importante relacionada a essa relação é raramente feita: qual é a amplitude possível ou permissível em R\P em uma estação ou ano? Um valor de R/P mínimo é fixado por custos de crescimento. O crescimento local pra a maioria das plantas superiores pode ocorrer com Y máximo de 0,80 – 0,85 mol C (mol C)-1, que é equivalente a um R/P mínimo de 0,15 – 0,20 mol de C (mol de C)-1. Quando os custos respiratórios da absorção de íons do solo, transporte ativo através o floema e assimilação de N são incluídos, o valor de R/P mínimo pode aumentar para cerca de 0,20 – 0,30. Por fim, é essencial alguma manutenção estrutural elevando os valores de R/P mínimo de longo prazo para cerca de 0,30 – 0,40 para a maior parte das plantas superiores . Dessa maneira, um decréscimo nos valores de R/P de 0,60 para 0,45 significa um grande aumento em crescimento por unidade de fotossíntese (sem mudança liquida na quantia de material de reserva) assim, variação aparentemente pequena em R/P pode ser significativa para o crescimento.
Estimativas para R/P de culturas são tipicamente menores do que pra vegetação natural. Valores relativamente pequenos de R/P em culturas poderiam estar relacionados com: uma grande fração de crescimento e biomassa das culturas está em órgãos de armazenamento tais como sementes e tubérculos comparado com uma pequena fração em outras plantas; Y em órgãos de armazenagem da maioria dos culturas de tubérculo e grãos é grande (0,83 –0,89) onde a respiração de crescimento é relativamente pequena; a respiração de manutenção nos órgãos de armazenamento é, em geral, lenta. Assim, ao selecionar genótipos para índice de colheita grande pode-se, indiretamente, selecionar para plantas com uma relação R/P menor de planta inteira.
Cálculos e medidas da respiração de crescimento.
A análise da rota de biossíntese permite estimar com bastante precisão o índice Y, partindo-se da composição de planta inteira ou de partes, desde que a eficiência real seja semelhante a eficiência potencial e o substrato seja conhecido. Ainda, deve-se atentar para as mudanças da composição da biomassa das plantas em seus vários estágios de desenvolvimento; por exemplo, a diferenciação e crescimento secundário podem ocorrer depois que órgãos são normalmente considerados maduros, em particular, a síntese de ligninas e hemiceluloses pode ser importantes em folhas após a expansão. E quando a ajustes fenotípicos ocorrem (como em folhas em resposta a mudança ambiental durante o desenvolvimento do dossel) a composição do tecido pode mudar, mudando o índice Y.
Assim, as medições de composição da biomassa usadas para calcular RG através de Y devem refletir quantidades de compostos sintetizados durante o crescimento (e não apenas acumulo liquido) para ser significativo. Além do que, a mobilização e senescência em órgãos velhos exige energia, mas não são explicadas em estimativas baseadas na rota de custos de crescimento; Penning de Vries et al (1983) destacou os custos de mobilização teóricos que podem ser especialmente importantes durante o enchimento de grãos em muitas culturas. Como exemplo da aplicação desse método, abordaremos a RG para a produção comercial de milho.
Além de estimar o RG a partir de estequiometrias de rotas bioquímicas e da composição da biomassa, outros métodos podem ser usados. Por exemplo, RG pode ser estimada derivando-se uma estimativa teórica ou experimental de RM e então subtraindo esse RM da respiração total medida (R).
1 – RG para a produção de café:
Partindo-se dos 5 principais compostos considerados por Penning de Vries et al. (1983), e do Y designado por esses autores para esses compostos a partir de 1 g de carboidrato fotossintetizado como sendo de: 0,404g de proteínas, 0,33g de lipídeos, 0,472g de lignina, 0,826g de carboidrato estrutural, 1,104g de ácidos orgânicos;considerando-se também a composição centesimal do grão de café torrado, podemos construir a seguinte tabela:
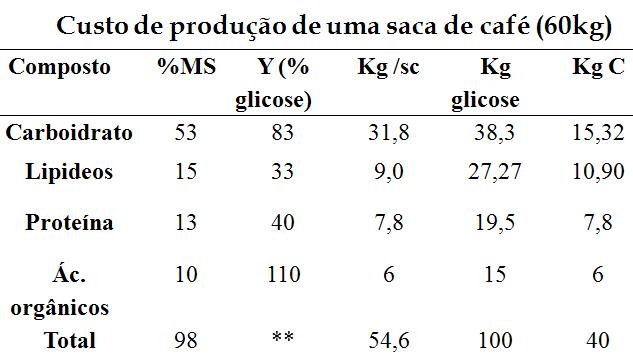
Onde: a %MS indica a quantidade de cada composto na semente de milho em percentagem, Y indica a eficiência de conversão de glicose em cada um dos compostos em termos de percentagem, kg/sc indica a quantidade de composto em uma saca de 60 kg de café, kg glicose indica o quanto essa quantidade (kg/sc) corresponde em glicose, para isso, usa-se o índice Y, kg C indica a quantidade de carbono, em massa, necessária para a síntese da referida quantidade de composto considerado, para isso, usa-se bases de peso molecular do carbono com base na glicose. Assim:
Y = ∆Ce/ ∆Sc
∆Ce = Kg substrato fixado em composto (54,6 kg/sc)
∆Sc = kg substrato utilizado para crescimento (Kg/glicose ou Kg/C)
Para 100 sacas de café / ha, podemos ter um consumo de 10.000 kg de glicose ou 4.000 kg de carbono, que são exportados nos grãos. O índice Y para o grão de café fica em torno de 0,54 (54,6 / 100), e indica que são necessários 1000 g de glicose para gerar um crescimento de 540 g de semente de milho. Com base em carbono, o Y = 1,36 (54,6 / 40), o que indica que são necessários 1,36 kg de carbono para produzir um crescimento de 1 kg de grão.
A partir desse exemplo, e conhecendo-se a composição de um órgão ou da planta inteira, pode-se estimar a sua respiração de crescimento, levando-se apenas em consideração as rotas de biossíntese dos compostos e o índice de eficiência de conversão Y.
Veja também:
Referências Bibliogáficas:
Buchanan, Gruissem, Jones, Biochemistry & Molecular Biology of Plants p. 696-705. 2000
Taiz, L.; Zeiger, E. Fisiologia vegetal. 5. ed. Porto Alegre: Artmed, 2012. 820 p.
Artigos: