
Nas plantas, o excesso de energia mitocondrial, acumulada sob a forma do gradiente eletroquímico de prótons, pode ser minimizado pela atuação da oxidase alternativa (AOX) (Fig. 1). A presença de grande número de caminhos não fosforilativos relacionados com o transporte de elétrons em mitocôndrias de plantas permite supor que estes caminhos estariam relacionados com o processo de adaptação das mesmas aos estresses ambientais.
A AOX foi o primeiro sistema dissipador de energia encontrado em plantas. Atualmente, tem sido demonstrado estar presente não somente em plantas superiores, mas também em fungos, leveduras e protozoários. Depende da atuação de uma proteína com massa molecular de aproximadamente 34 kDa, caracterizada como sendo uma enzima dímera, cujas subunidades são ligadas por ponte dissulfídica. A AOX é sensível ao ácido salicil-hidroxâmico (SHAM) e ao ácido benzohidroxâmico (BHAM). Em alguns tecidos, pode ser estimulada por ácidos, como o piruvato. A AOX é um dímero em sua forma inativa (oxidada). O dímero é dividido em duas proteínas para ficar em sua forma ativa (reduzida). Para o propósito de transportar os elétrons, a AOX tem um sítio de ligação putativa de ferro em seu C terminal e a ponte de dissulfeto entre duas moléculas de cisteína o qual possibilita a formação das formas ativa e inativa está localizado em seu N terminal.
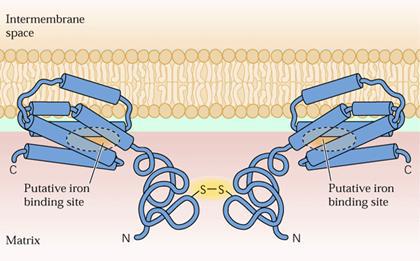
Figura 1. Conformação da oxidase alternativa
A AOX é caracterizada como participante de um caminho não fosforilativo em plantas e pode ter papel importante em plantas termogênicas da família Araceae, nas quais o aumento da temperatura promove a volatilização de aminas, cuja função está relacionada com a atração de insetos polinizadores . Aumentos de temperatura, também são observados em frutos climatéricos em estádio de amadurecimento e isso, algumas vezes, tem sido atribuído à atividade da AOX (Fig. 2).
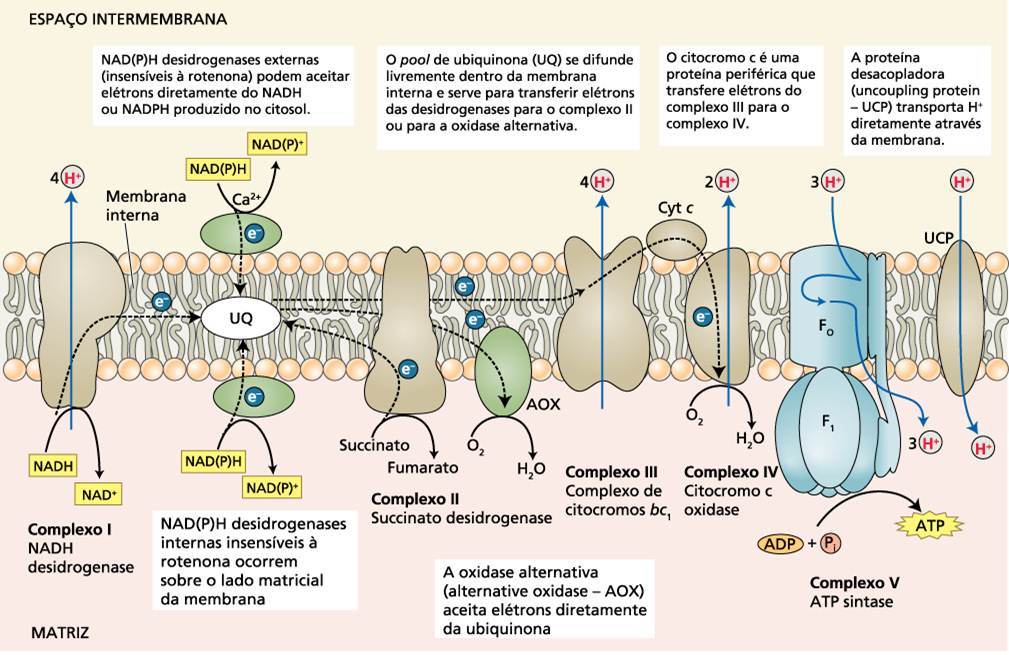
Figura 2. Cadeia transporte de elétrons com a oxidase alternativa.Fonte: Taiz (2013)
Em plantas não termogênicas, a AOX possui papel fundamental no que se refere ao balanço energético da célula. Além disso, vários estudos independentes propõem ter a AOX capacidade de prevenir a produção de ROS, proveniente de várias formas de estresses biótico e abiótico. Por exemplo, em trigo “durum”, a AOX pode atuar como sistema dissipador de energia,podendo controlar a geração de ROS. Experimentos realizados com suspensões celulares de tabaco transgênico, com super-expressão da AOX, mostraram diminuição na quantidade de ROS após a adição de antimicina A . Outros experimentos têm mostrado que o estresse pelo frio é responsável pela ativação da AOX em batatas e outras plantas.
Outra forma de dissipação do excesso de energia mitocondrial envolve a participação da proteína desacopladora de mitocôndrias de plantas (PUMP).Essa proteína age de modo semelhante à ação das proteínas desacopladoras presentes em mitocôndrias de tecidos animais (UCP), dissipando o gradiente de prótons da membrana mitocondrial, na presença de ácidos graxos livres, sendo fisiologicamente inibida por ATP, GDP e GTP . A PUMP foi caracterizada como uma proteína de massa molecular em torno de 32 KDa, cuja função inicial supunha-se ser apenas termogênica. Posteriormente, foi verificado o seu envolvimento no amadurecimento e senescência de frutos de tomate. A PUMP tem sido identificada por imunodetecção, em vários órgãos e tecidos de mais ou menos 30 espécies de plantas, incluindo monocotiledôneas e dicotiledôneas; C3, C4 e plantas CAM; frutos climatéricos e não climatéricos; bem como em tecidos termogênicos e não termogênicos.
Muitas plantas floridas atraem insetos polinizadores liberando moléculas odoríferas que mimetizam uma fonte de alimento natural de insetos ou locais potenciais de postura de ovos. Algumas plantas polinizadas por moscas ou escaravelhos que normalmente se alimentam ou põem seus ovos em esterco ou carne podre usam compostos com malchei para atrair esses insetos.
Uma família de plantas malcheirosas é a Araceae, que inclui os filodendros, os copos-de-leite e de uma espécie de couve malcheirosa. Essas plantas apresentam flores diminutas densamente empacotadas em uma estrutura ereta chamada espádice, rodeada por uma folha modificada de- nominada espata. A espádice libera odores de carne putreata ou esterco. Antes da polinização, a espádice também se aquece, em muitas espécies, de 20 até 40°C acima da temperatura ambiente. A produção de calor (termogênese) ajuda a evaporação das moléculas odoríferas para uma melhor dispersão. Como a carne putrefata e o esterco são geralmente quentes, devido ao metabolismo hiperativo dos micróbios que deles se alimentam, o próprio calor também pode atrair insetos. No caso da couve oriental malcheirosa, que floresce no final do inverno ou no começo da primavera, quando a neve ainda cobre o solo, a termogênese permite que a espádice cresça através da neve.
As mitocôndrias das plantas, fungos e eucariotos unicelulares apresentam sistemas de transporte de elétrons que são essencialmente os mesmos que os dos animais, mas elas também possuem uma via respiratória alternativa. Nesta via, uma QH2 oxidase resistente ao cianeto transfere os elétrons do reservatório de ubiquinona diretamente para o oxigênio, desviando-os das duas vias de translocação de prótons dos complexos III e IV . A energia que poderia ser conservada como ATP é liberada como calor. A mitocôndria das plantas também apresenta uma desidrogenase do NADH alternativa que é insensível à rotenona, uma substância inibidora do complexo I, que transfere elétrons do NADH na matriz diretamente para a ubiquinona, desviando-os do complexo I e seu bombeamento associado de prótons. As mitocôndrias das plantas apresentam ainda uma outra desidrogenase do NADH, na face externa da membrana interna, que fica defronte ao espaço intermembranas e transfere os elétrons do NADPH ou NADH para a ubiquinona, desviando-os novamente do complexo I. Assim, quando os elétrons entram na via respiratória alternativa através da desidrogenase do NADH insensível à rotenona, a desidrogenase do NADH externa ou desidrogenase do succinato (complexo II), e passam para o O2 via oxidase alternativa resistente ao cianeto, a energia não é conservada como ATP, mas liberada como calor. A couve malcheirosa pode usar o calor para derreter a neve, produzir calor de podre ou atrair escaravelhos ou moscas.
Principais funções:
- Controle de pH
Um pressuposto discutido por Sakano (1998) é o controle de pH, desde que o complexo I transporta prótons para o espaço intermembranas, então o pH depende também da atividade do complexo I. Considerando que a AOX aumenta atividade adicional do complexo I, o pressuposto do controle de pH é suportado. Numa condição de excesso de prótons, a respiração alternativa pode facilitar a eliminação destes prótons indesejáveis, de maneira menos limitada do que a respiração citocrômica.
- Compensação para a Inibição por CN–:
Outro pressuposto generalista é que a AOX é resistente aos inibidores da respiração citocrômica, isto pode parcialmente compensar para os danos feitos por eles. A respiração alternativa contribui para o gradiente de prótons, e portanto para a síntese de ATP. O assunto é discutido por Milenaar et al. (2002), onde a menor sensibilidade ao cianeto é mencionada como resultado do aumento da atividade da AOX devido ao citrato. Esse pressuposto é muito limitado, desde que ele pode somente estar certo para quantidades muito pequenas de CN–.
- Eliminação de EROs:
A eliminação de EROs, está relacionada com a absorção de fosfato. Esse aspecto é revisado em Juszczuk et al. (2001) e em um extenso artigo bem como por Maxwell et al. (1999). As EROs são as formas iniciais das espécies Ativas de Oxigênio ou radicais livres. Elas apresentam características tóxicas, e os organismos desenvolveram mecanismos para eliminar as EROs. Estatisticamente, as EROs estão presentes entre as moléculas de oxigênio não ativas. Quando muito oxigênio está presente, a probabilidade é alta de existirem EROs. As EROs moléculas pode ser eliminadas por reação química com dois prótons e dois elétrons para produzir H2O. A razão para a entrada de oxigênio nas raízes é a absorção de fosfato. O Fosfato é um dos mais importantes nutrientes minerais para os organismos e ele pode ser um fator limitante para o crescimento dos organismos. Em um experimento com raízes de feijão, foi observado que a absorção de oxigênio é fortemente correlacionada com absorção de fosfato A absorção de fosfato aumenta quando há uma condição de stress por carência de fosfato. A planta precisa investir mais esforço para obter a quantidade de fosfato que ela precisa. E então como dito anteriormente, a absorção de oxigênio é aumentada e as EROs estão provavelmente presentes em maior quantidade.
- Viabilidade:
Além da eliminação das EROs, em botânica, a melhor maneira de descobrir sobre as vantagens de um certo componente é a supressão dele. A supressão da AOX é realizada pelo AS8, o qual é o anti-sense da AOX, o mutante não apresenta atividade da oxidase alternativa. Pode-se observar na tabela que a taxas de viabilidade do tipo selvagem são elevadas em relação ao mutante e que o peso seco do mutante é alto em relação ao tipo selvagem. O pressuposto é que a AOX aumenta a taxa de sobrevivência do tipo selvagem, e que sob stress de fosfato a estratégia de sobrevivência seria a limitação da taxa de crescimento, pois na cultura de células a massa é menor na cultura do tipo selvagem, ou seja, o tipo selvagem cresce menos conseguindo sobreviver com uma quantidade limitada de fósforo.
- Termogênese:
Buchanan et al (2000) discutem uma função muito interessante para a respiração alternativa. A termogênese (aumento de temperatura) é realizada pela AOX. A este respeito seria apropriado distinguir os órgãos florais termogênicos em relação às plantas que não apresentam esse tipo de estrutura. Os lírios da água apresentam órgãos florais termogênicos. Estes órgãos florais termogênicos podem elevar sua temperatura em 10°C acima da temperatura ambiente.Nem toda a energia disponível é usada para aumentar o gradiente de prótons, e o excesso de energia é liberado como calor. A razão para que as plantas termogênicas elevem a temperatura de seus órgãos florais a taxas tão altas deve-se à presença de muito mais mitocôndrias nos órgãos termogênicos. A finalidade da termogênese nas plantas é a volatilização de compostos químicos e para atrair polinizadores ou pode mimetizar o odor de carniça para atrair determinados insetos que depositam seus ovos nas flores.
Veja também:
Referências Bibliogáficas:
Buchanan, Gruissem, Jones, Biochemistry & Molecular Biology of Plants p. 696-705. 2000
Taiz, L.; Zeiger, E. Fisiologia vegetal. 5. ed. Porto Alegre: Artmed, 2012. 820 p.
Artigos: