
A fotossíntese ocorre na organela subcelular conhecida como cloroplasto. Nele, existe um extenso sistema de membranas internas conhecidas como tilacóides, onde toda a clorofila está contida e onde ocorrem reações ligadas à etapa fotoquímica. Já as reações de fixação de carbono ocorrem no estroma, região que circunda os tilacóides e onde estão contidas enzimas hidrossolúveis (Figura 1).
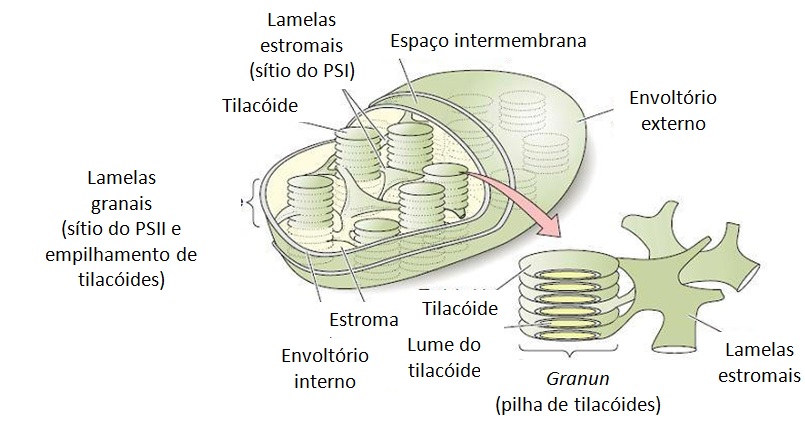
Figura 1. Organização geral das membranas do cloroplasto (TAIZ; ZEIGER, 2004).
Nos cloroplastos ocorre a reação da mais fundamental importância para a vida das plantas e, indiretamente, para a vida dos animais, a fotossíntese. Os cloroplastos são geralmente discoidais e sua cor é verde devido à presença de um pigmento denominado clorofila. No seu interior existe um conjunto bem organizado de membranas, as quais formam pilhas unidas entre si, que são chamadas de grana. Cada elemento da pilha, que tem o formato de uma moeda, é chamado de tilacóide. Já o estroma (fluído gelatinoso que preenche o cloroplasto) existem enzimas, DNA, pequenos ribossomos e amido. As moléculas de clorofila se localizam nos tilacóides, reunidas em grupos.
No cloroplasto, a energia luminosa é colhida pelos fotossistemas e é usada para permitir a transferência de elétrons por uma série de compostos que agem como doadores de elétron e receptores de elétron. A maioria de elétrons ao final do processo, reduz NADP+ em NADPH. A energia luminosa também é usada para gerar uma força motiva de próton através da membrana do tilacóide, usada para sintetizar ATP.
Ontogênese
Os cloroplastos são formados a partir de fissão binária de cloroplastos jovens ou completamente maduros preexistentes, independentemente da divisão celular. Porém, em tecidos meristemáticos, onde só existem proplastídeos (estágio anterior aos cloroplastos), a divisão da organela ocorre juntamente com a divisão celular, sendo que cada célula filha possui aproximadamente o mesmo número de plastídeos que a célula mãe. Entretanto, na fase de expansão celular, eles continuam se dividindo. Semelhantemente, a replicação do DNA das organelas não está limitada à fase S do ciclo celular, quando o DNA nuclear se replica, mas ocorre ao longo de todo o ciclo. As moléculas individuais de DNA das organelas parecem ser aleatoriamente selecionadas para a replicação, de tal forma que, em um dado ciclo celular, algumas podem se replicar mais de uma vez e outras simplesmente não se replicam.
Os proplastídeos são plastos pequenos, incolores, não diferenciados, limitados por uma dupla membrana, abundantes em células meristemáticas e estão ausentes nas membranas internas. Sua membrana interna é pobremente desenvolvida, consistindo de poucas invaginações e um pequeno número de vesículas. Na presença da luz, a membrana interna cresce e emite vesículas que são liberadas para a matriz e são transformadas em discos, que vão formar os tilacóides. No cloroplasto maduro, os tilacóides não estão mais conectados à membrana interna, porém os grana permanecem unidos por tilacóides intergrana (Figura 2).
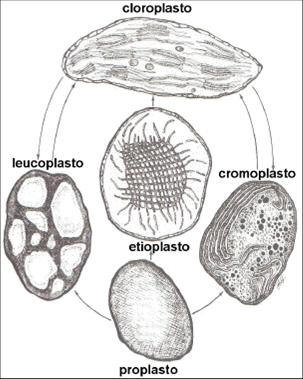 Figura 2. Ontogênese de cloroplastos.
Figura 2. Ontogênese de cloroplastos.
Caso a planta seja colocada em baixa intensidade luminosa ou crescer desde o início com pouca intensidade de luz, o processo ocorre em ordem, provocando a desorganização das membranas, fenômeno este chamado de estiolamento. Nesse caso, as vesículas do proplastídeo agregam-se para formar um ou vários corpos prolamelares, os quais podem desenvolver-se em grana quando expostos à luz. Esses compartimentos formados a partir de cloroplastos, devido à ausência ou pouca luz, denominam-se estioplastos, que apesar de não possuírem clorofila, produzem uma grande quantidade de seus precursores, os protoclorofilídeos, e apresentam lipídeos de membrana na forma de corpos prolamelares. Quando iluminados, começam a rediferenciar-se em cloroplastos. Isso mostra a marcante capacidade de diferenciar-se, desdiferenciar-se e rediferenciar-se .
Os proplastídeos são os precursores de todos os membros da família plastídeo. Etioplasto é um estágio transitório entre os proplastídeos iluminados e os cloroplastídeos e são abundantes em folhas estioladas. O desenvolvimento dos cloroplastídeos ocorre simultaneamente com o enverdecimento das plantas, resultado da síntese de clorofilas. Os cromoplastos são plastídeos pigmentados de coloração amarelada a alaranjada, em função da presença de carotenóides os quais não apresentam habilidade para realizarem a fotossíntese. Os amiloplastídeos estruturas especializadas na síntese e armazenamento de amido, sendo, portanto, incolores. O termo leucoplastos, muito comum na literatura não se refere a nenhum tipo especial de plastídeo e, sim, a todos os plastídeos não pigmentados.
As proteínas do cloroplasto podem ser codificadas tanto pelo DNA do cloroplasto quanto pelo do núcleo. As proteínas codificadas no cloroplasto são sintetizadas em ribossomos do cloroplasto, enquanto que aquelas codificadas no núcleo o são em ribossomos citoplasmáticos e posteriormente transportadas para os cloroplastos.
Os genes requeridos para a codificação do funcionamento do cloroplasto estão distribuídos no núcleo e no genoma do cloroplastídeo. Esses genes não possuem um padrão definido, porém ambos os conjuntos são essenciais para manutenção da viabilidade do cloroplasto. Alguns genes do cloroplasto são necessários para outras funções celulares, tais como a síntese de lipídeos. O controle da expressão dos genes nucleares que codificam para as proteínas do cloroplasto é complexo, envolvendo regulação dependente da luz, mediada pelo fitocromo e pela luz azul, bem como outros fatores.
O número de proteínas codificadas pelo genoma da organela é pequeno, e mesmo assim os cloroplastos conduzem sua própria replicação e transcrição do DNA e síntese protéica. Esses processos ocorrem no estroma.
Embora as proteínas que medeiam tais processos genéticos sejam particulares às organelas, a maior parte delas é codificada pelo genoma nuclear.
As poucas proteínas codificadas pelos genomas destas organelas estão localizadas, em sua maioria, na membrana do tilacóide. Estes polipeptídios codificados pela organela geralmente são subunidades de complexos protéicos, cujas demais subunidades são codificadas por genes nucleares e são importadas do citosol.
Em mutantes, apesar de algumas células serem incapazes de processar a síntese cloroplastídica de proteínas, ainda assim elas apresentam cloroplastos e tornam-se perfeitamente viáveis, caso lhes sejam fornecidos substratos oxidáveis. Se o amadurecimento dos cloroplastos é bloqueado em vegetais superiores, entretanto, seja pelo crescimento das plantas no escuro ou porque o DNA cloroplastídico apresenta-se defeituoso ou ausente, estes vegetais morrem assim que os seus estoques de nutrientes terminam.
Uma vez que a maioria dos genes codificantes das atuais proteínas cloroplastídicas está no núcleo celular, parece claro que uma extensa transferência de genes ocorreu do DNA das organelas para o DNA nuclear durante a evolução eucariótica. Isso explicaria porque alguns dos genes nucleares codificantes de proteínas cloroplastídicas assemelham-se a genes bacterianos.
Diferenças tecido-específicas são encontradas entre as proteínas de cloroplastos codificadas pelo núcleo. Isso quer dizer que os cloroplastos podem ter funções especializadas em tipos particulares de células.
As proteínas sintetizadas no citosol, por polirribossomos, são traduzidas com um peptídeo sinal na extremidade N-terminal, de 40 a 50 aminoácidos, que as direciona ao cloroplasto e capacita a sua translocação através das membranas. O processo de importação de proteínas requer chaperoninas dentro e fora do cloroplasto e um grupo de proteínas chamadas aparato de importação de proteínas, que promovem a união entre a membrana externa e interna do cloroplasto no ponto de entrada da proteína importada. Ambos os passos requerem energia (ATP). As chaperoninas citosólicas se ligam à proteína que está sendo sintetizada, impedindo que ela atinja sua configuração nativa. Então, os peptídeos em transição interagem com os lipídeos da membrana externa ou com o aparato de importação de proteínas, o qual forma um canal proteináceo permitindo a passagem do polipeptídeo para dentro da organela. Em um segundo passo, o peptídeo sinal é removido por proteases do estroma. Se a proteína pertence ao estroma, outras chaperoninas ligam-se a ela, conferindo-lhe a conformação final.
As chaperoninas são proteínas citosólicas pertencentes à família das hsp70. Tanto auxiliam para garantir a conformação correta de proteínas citosólicas, quanto desempenham uma função essencial na importação de proteínas para as mitocôndrias e para o retículo endoplasmático rugoso se ligando a proteína precursora, sem sua conformação nativa, durante o transporte. A liberação do polipeptídeo recentemente sintetizado para as chaperoninas citosólicas necessita da hidrólise de ATP.
Acredita-se que as proteínas precursoras recém-sintetizadas liguem-se às proteínas hsp70 assim que são liberadas dos polirribossomos no citosol, evitando que estas se agreguem ou assumam espontaneamente sua conformação final antes de se ligarem à proteína transportadora na membrana da organela de destino. A energia liberada pela hidrólise de ATP é usada para liberar as hsp70 ligadas assim que a proteína passa através da membrana.
Uma vez que se encontram no interior da organela, as proteínas são recebidas na matriz por proteínas auxiliares muito relacionadas com as hsp70. As proteínas hsp70 da organela se ligam firmemente a proteína transportada assim que esta é liberada da proteína transportadora. A hsp70 libera a proteína em um passo dependente de ATP. Este ciclo de ligação e subseqüente liberação, movido à energia, propicia a força para importação da proteína após sua inserção inicial na proteína transportadora. A ligação subseqüente de múltiplas hsp70 da organela pode impulsionar a proteína através do canal transmembrana formado pelo aparato de importação de proteínas.
Depois da interação inicial com as hsp70, as proteínas importadas que pertencem ao estroma são transferidas para outra chaperonina, a hsp60. Esta se liga à cadeia polipeptídica desdobrada e facilita a transição para a conformação final da proteína em uma reação que consome ATP.
Os cloroplastos, que possuem um gradiente de potencial eletroquímico através da membrana do tilacóide, mas não na sua membrana interna, parecem empregar apenas a hidrólise do ATP como força para importar proteínas através do seu envelope externo de dupla-membrana.
Se a proteína é destinada ao lúmen do tilacóide, ela possui um segundo peptídeo sinal que a guia através da membrana do tilacóide. Assim, chaperoninas cloroplastídicas se ligam ao polipeptídeo durante seu transporte até a membrana do tilacóide. No lúmen, uma outra protease separa o peptídeo sinal da proteína.
Muitas proteínas do cloroplasto, incluindo subunidades do sistema fotossintético e da ATP sintetase, estão embebidas na membrana dos tilacóides. Estas proteínas são transportadas do citosol até seu destino em duas etapas. Primeiro, elas atravessam a membrana dupla em sítios de contato até o estroma, e, então, são transportadas até a membrana tilacóide (ou através desta membrana até o interior do tilacóide). Os precursores dessas proteínas possuem um peptídeo-sinal tilacóide hidrofóbico, após o peptídeo-sinal amino-terminal do cloroplasto. Depois que o peptídeo-sinal amino-terminal é usado para importar a proteína até o estroma, ele é removido por uma enzima sinal-peptidase do estroma (protease). Esta clivagem expõe o peptídeo-sinal do tilacóide, que então inicia o transporte através da membrana do tilacóide. A segunda etapa é a via usada para inserir proteínas codificadas pelo cloroplasto na membrana tilacóide. A proteína transportadora necessária, presumivelmente, é originária da bactéria ancestral do cloroplasto.