
Determinação do estado hídrico de tecidos vegetais (Teor de Água; Teor Relativo de Água; Potencial Hídrico) (Baixa o arquivo em PDF)
ATIVIDADE PRÁTICA
DETERMINAÇÃO DO ESTADO HÍDRICO EM TECIDOS VEGETAIS (Teor de Água, Teor Relativo de Água e Potencial Hídrico
OBJETIVOS:
- Ter contato com as possibilidades de ter contato com os principais métodos que podem ser utilizados para determinar/estimar o estado hidrico de uma célula, tecido, orgão de uma planta;
- Vantagens/desvantagens de cada método e identificação das principais dificuldades e cuidados na utilização dos mesmos
- Consolidar o entendimento das bases termodinamicas e biofisicas que envolve as relações hidricas num sistema solo-planta-atmosfera.
INTRODUÇÃO
A água está presente em todas as partes da planta. Cada célula vegetal está envolvida ou impregnada com água, sendo portanto, a molécula em atividade mais abundante nas células e órgãos da planta, constituindo-se, quase sempre, em mais de 80% do peso da matéria vegetal fresca. Além de exercer atividades físicas como a ação mecânica no alongamento celular, também atua como reagente em inúmeras reações metabólicas.
Devido à sua importância, o conhecimento de técnicas para as medições de seu teor nos tecidos torna-se necessário. Existem várias terminologias e conceitos para caracterizar o estado hídrico de uma célula, tecido ou órgão vegetal. Usa-se caracterizar o estado hídrico utilizando-se do potencial da água (ya) e seus componentes (yp e ys ) e também do teor de água. Pretende-se neste estudo, mostrar alguns métodos de determinação e caracterização das condições hídricas dos vegetais.
A – DETERMINAÇÃO DO TEOR DE ÁGUA (TA)
Usa-se a seguinte equação para se calcular o teor de água (TA) com base nos pesos do material vegetal fresco (Pf) e seco (Ps) numa temperatura variando de 60 a 70°C.
TA = (Pf – Ps) / Pf x 100
Caso seja impossível secar o material logo após a coleta, convém pesá-lo imediatamente para se obter o peso fresco sem subestimá-lo.
B – DETERMINAÇÃO DO TEOR RELATIVO DE ÁGUA (TEOR RELATIVO E DÉFICIT DE SATURAÇÃO DE ÁGUA)
O teor relativo de água (TRA) pode ser calculado utilizando-se a seguinte equação:
TRA = (Pf – Ps) / Pst – Ps) x 100
O termo déficit de saturação de água (DSA) pode ser definido como a quantidade de água que a planta ou parte dela requer para atingir a saturação (100%). Daí nota-se que os termos DSA e TRA são complementares, sendo TRA = 100 – DSA.
Para obtenção do peso saturado (Pst), coloca-se o material sobrenadando em água destilada ou deionizada até atingir peso constante. Cuidar para que o tecido não perca ou ganhe material por respiração ou fotossíntese. Para tanto, utiliza-se técnicas que inabilitem a continuação das atividades fisiológicas.
Para obtenção do Pst de folhas pode-se utilizar discos foliares que atingem a saturação normalmente no tempo de 4 a 6 horas. Este tempo pode ser determinado experimentalmente relacionando-o com o aumento do peso dos discos, até peso constante. Os discos após a saturação e antes da pesagem têm suas superfícies secadas em papel de filtro sempre de maneira semelhante: mesma pressão, mesma pessoa, etc. tentando diminuir os erros decorrentes da operação.
Pode-se utilizar de 8 a 15 discos foliares com 0.5 a 2.0 cm de diâmetro, em pelo menos 3 repetições, que se obtém bons resultados. Depois de determinado o peso saturado (Pst), obtém-se o peso seco em estufa a 70 – 80°C e normalmente após 4 horas as amostras atingem peso constante.
Todo o cuidado é pouco na determinação do TRA, pois representa com muita fidelidade o nível da turgescência celular, lembrando que um tecido vegetal sem déficit hídrico o valor de TRA é de 100%
QUESTIONÁRIO:
- Cite os principais cuidados necessários na obtenção do peso fresco, peso de saturação, componentes da equação de determinação do TRA.
- Que erro(s) seria(m) provocado(s) na determinação do teor de água pela demora na determinação do peso fresco?
C – ESTIMATIVA DO POTENCIAL HÍDRICO DE TECIDOS VEGETAIS PELO MÉTODO DA BOMBA DE PRESSÃO (SCHOLANDER)
INTRODUÇÃO
Considerando que os estudos de relações hídricas baseiam-se nos princípios de movimentação da água no sistema solo-planta-atmosfera, é necessário que se conheça o significado de potencial hídrico (representado pelo símbolo yw e seus componentes, que são:
No solo: ψa =ψp + ψg + ψs + ψm
Na planta: ψa = ψs + ψp
Onde,
ψw = Potencial total da água, representa a energia livre da água.
ψp = Potencial de pressão, cujo valor mínimo é zero e assume valores cada vez maiores à medida que aumenta a pressão provocada por algum peso sobre um dado ponto. Por exemplo, na célula é a pressão exercida pela água no seu interior contra a parede celular. Logo, quanto mais túrgida estiver a célula, maior é o ψp.
ψg = Potencial gravitacional, que aparece devido a existência do campo gravitacional terrestre e assume valores negativos uma vez que a ação da gravidade contribui para diminuir a energia livre da água. Considerado desprezível na planta.
ψs = Potencial de soluto ou osmótico, que corresponde à concentração de sais na solução do solo ou do suco celular. Seus valores são sempre negativos, uma vez que aumentando a concentração de solutos numa solução diminui-se a energia livre da água.
Lembre que ψs= – MRTi (M=concentração molar; R=constantes dos gases; T=temperatura em kelvin; i= constante de ionização)
ψ m = Potencial mátrico, representa a água adsorvida nas partículas sólidas do solo. Na planta é considerado desprezível, uma vez que a fração de água adsorvida na parede celular é irrisória e desprezível. Apresenta valores sempre negativos.
Numa célula vegetal adulta a maior parte do volume é ocupado por um ou mais vacúolos grandes, cheio (s) de solução aquosa rica em substâncias orgânicas e inorgânicas osmoticamente ativas, o que capacita à célula absorver água. No início, a célula estando flácida (murcha), a pressão interna exercida contra a parede celular é insignificante e, nesse caso, considera-se que ψp = 0. À medida que vai entrando água no seu interior, alguma pressão começa a existir contra a parede celular e, portanto, o potencial de pressão (ψp) assume valores cada vez maiores e positivos, até a célula atingir turgidez total (máxima capacidade de retenção de água), quando o potencial hídrico da célula (ψw) é zero e o potencial de pressão (ψp) é máximo. À medida que a célula perde água a concentração de solutos é aumentada no seu interior e o potencial de soluto ou osmótico (ψs) torna-se cada vez mais negativo. Na natureza o permanente acúmulo de solutos no suco vacuolar provenientes da fotossíntese e absorção de íons do solo resulta num abaixamento do potencial osmótico.
“Não podemos ignorar que o aumento na concentração de Solutos Osmoticamente Ativos (S.O.A), ou seja a redução no potencial de soluto ou osmótico (ys) de uma célula, pode ser decorrente da redução no volume da agua (solvente) celular ou no aumento de moléculas de SOA nas células, decorrentes de ações estratégicas conhecidas por ajustamento osmótico celular”
METODOLOGIA
Neste método conhecido como “método da bomba de pressão” ou de SCHOLANDER et al (1965), uma folha, após destacada da planta, é imediatamente colocada num cilindro de maneira que apenas o pecíolo fique para fora, atravessando um pequeno orifício revestido de borracha na tampa (à prova de gás). Uma pressão é gradativamente fornecida à câmara do cilindro até que se verifique o primeiro sinal de expulsão da seiva na extremidade exposta do pecíolo, quando a injeção do gás (inerte-nitrogênio) deve ser interrompida e a leitura verificada num manômetro. O valor encontrado representa o potencial de pressão do xilema, onde o potencial de soluto ou osmótico é considerado desprezível. Neste caso, admite-se que yw = yp. Expressa-se os valores de ya em MPa (Mega Pascal). 1 MPa = 10 atm.
Fotografias e representação esquemática de câmaras de pressão usadas para medir o potencial de água (Ψa) de uma folha ou ramo com folhas (https://en.wikipedia.org/wiki/Scholander_pressure_bomb#cite_note-1)


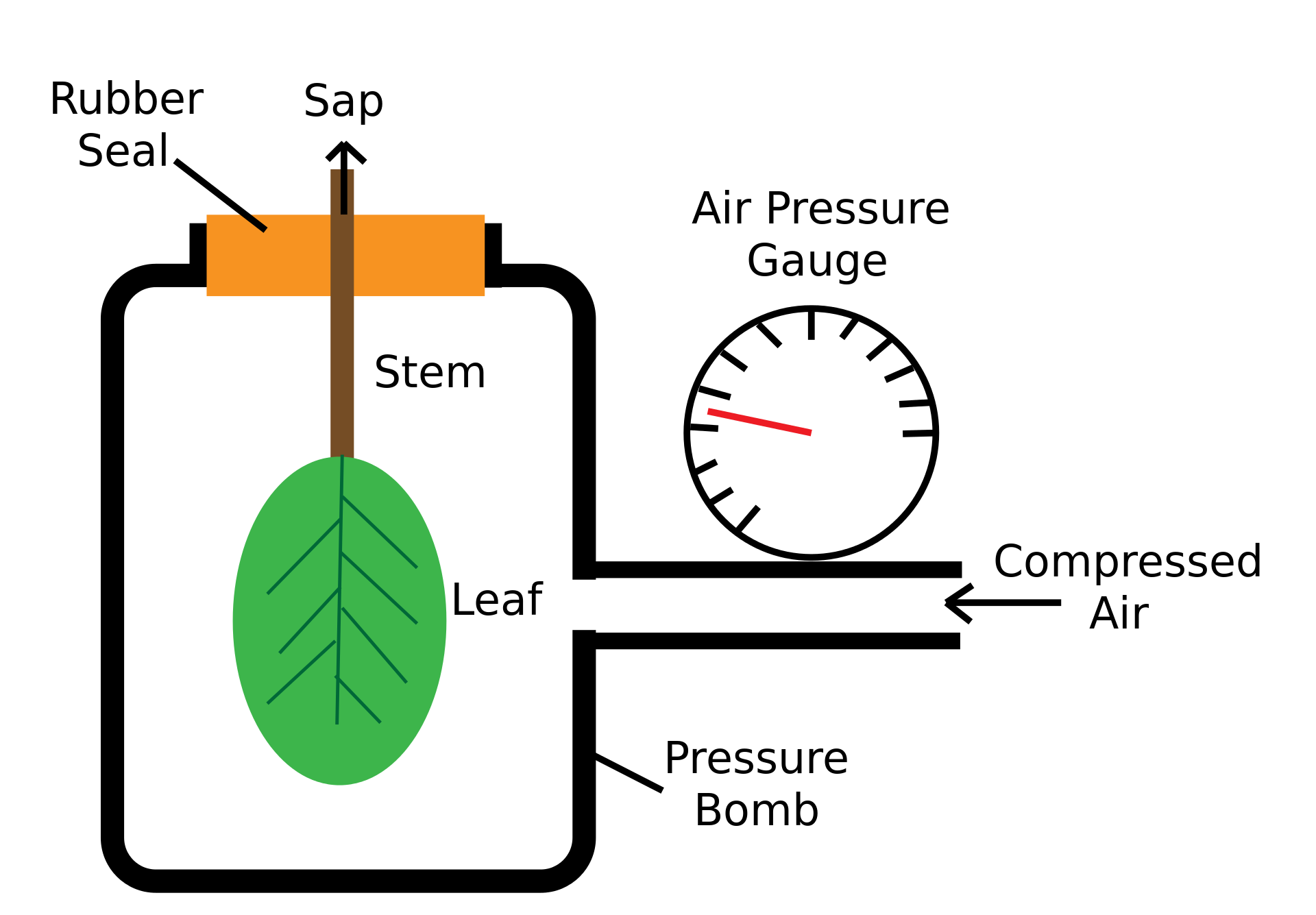
Uma bomba de pressão ou câmara de pressão ou bomba Scholander é um instrumento que pode medir o potencial aproximado de água dos tecidos vegetais. Um segmento de folha e pecíolo ou caule é colocado dentro de uma câmara selada. Gás pressurizado (nitrogênio normalmente comprimido) é adicionado lentamente à câmara. À medida que a pressão aumenta em algum momento, o conteúdo líquido da amostra será forçado a sair do xilema e ficará visível na extremidade cortada da haste ou pecíolo. A pressão necessária para isso é igual e oposta ao potencial hídrico da amostra (folha ou total). As bombas de pressão são portáveis em campo e mecanicamente simples, o que as torna o método predominante para medições de potencial hídrico nos campos da fisiologia e ecofisiologia das plantas. Várias variáveis de potencial hídrico podem ser determinadas usando a análise de bomba de pressão. Os mais comuns são o potencial de água nas folhas antes do amanhecer e o potencial de água nas folhas ao meio-dia. As medições realizadas nas plantas antes do amanhecer são consideradas uma boa representação do estado hídrico da planta. Como nenhuma transpiração através dos estômatos deve ocorrer à noite, o potencial hídrico da planta deve estar em equilíbrio em toda a planta e ser semelhante ao potencial hídrico do solo ao redor das raízes. O potencial hídrico foliar ao meio-dia é menos utilizado, e usado com cuidado, pois é mais variável e não se correlaciona bem com outras medidas fisiológicas do estado da água. No entanto, os potenciais hídricos ao meio-dia podem ser usados para determinar os períodos de pico de deficit/estresse hídrico ou alterações diurnas no estado hídrico da planta. [5] Variáveis e métodos adicionais que envolvem bombas de pressão para análise incluem: condutância da haste, embolias do xilema, etc.
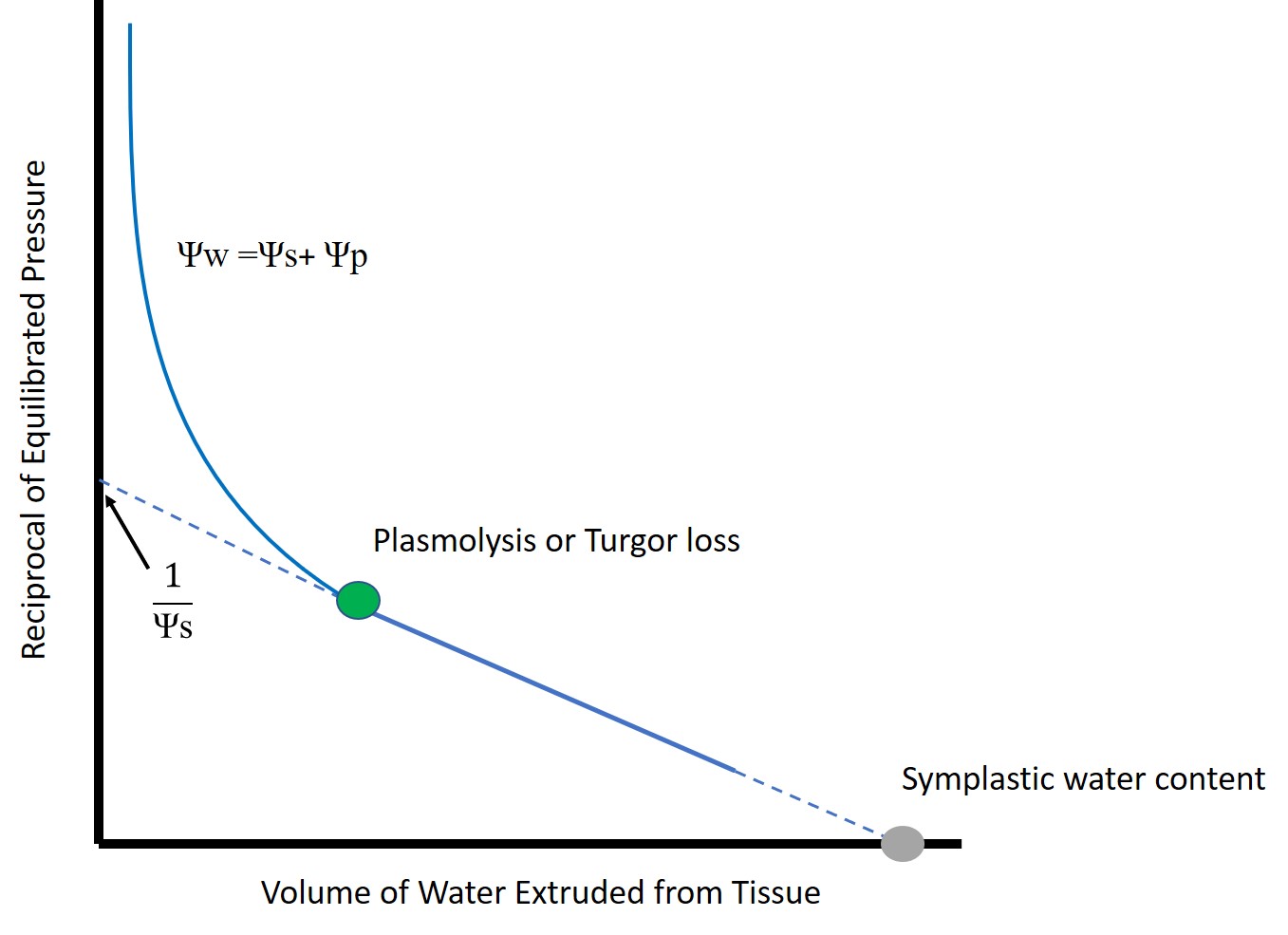
O uso de Curvas pressão-volume utilizando a Bomba de Pressão e TRA(Faça leitura do texto acompanhando a figura simplificada postada a seguir que facilitará a compreensão)
Um método mais avançado que utiliza a bomba de pressão na fisiologia das plantas é a análise de curvas pressão-volume ou curva p-v. Através desse método, mede-se as alterações no potencial hídrico (Ψa) das folhas ou caule e no teor relativo de água (TRA) para isolar os componentes subjacentes do potencial total de água nas folhas ou caule. Embora as medições possam demorar muito tempo, variáveis como potencial de soluto (Ψs), ponto de perda de turgor (Ψtlp), teor de água apoplástica e teor de água simplástica podem ser determinadas usando esse método. O protocolo geral para medir curvas p-v envolve medidas repetidas de potencial e massa de água em sucessão. Como a água é forçada a sair da amostra a cada medição na bomba de pressão, a massa também é reduzida. O rastreamento dessas alterações em muitas medições deve mostrar uma queda precipitada e, em seguida, um declínio linear constante após um ponto de inflexão
Figura mostrando a Curva pressão-volume simplificada
Referencias sugeridas para leitura que poderão auxiliar num maior aprofundamento dos tema abordados e sugeridaos na Wikipedia
- http://jxb.oxfordjournals.org/cgi/content/abstract/23/1/267 1972 Journal of Experimental Botany – The Measurement of the Turgor Pressure and the Water Relations of Plants by the Pressure-bomb Technique M. T. TYREE and H. T. HAMMEL
- Scholander, P.; Bradstreet, E.; Hemmingsen, E.; Hammel, H. (1965). “Sap Pressure in Vascular Plants: Negative hydrostatic pressure can be measured in plants”. Science. 148 (3668): 339–346. Bibcode:..148..339S. doi:10.1126/science.148.3668.339. PMID17832103.
- Gifftord, H.H. (18 August 1971). “SIMPLIFIED APPARATUS FOR DETERMINING LEAF WATER POTENTIALS IN PINE NEEDLES” (PDF). Scion Research.
- Améglio, Thierry; Archer, Philippe; Cohen, Moisés; Valancogne, Charles; Daudet, François-alain; Dayau, Sylvia; Cruiziat, Pierre (1999). “Significance and limits in the use of predawn leaf water potential for tree irrigation”. Plant and Soil. 207 (2): 155–167. doi:1023/A:1026415302759.
- Williams, L.E.; Araujo, F.J. (2002). “Correlations among Predawn Leaf, Midday Leaf, and Midday Stem Water Potential and their Correlations with other Measures of Soil and Plant Water Status in Vitis vinifera”. Journal of the American Society for Horticultural Science. 127 (3): 448–454. doi:21273/JASHS.127.3.448.
- Cochard, Hervé; Cruiziat, Pierre; Tyree, Melvin T. (1992). “Use of Positive Pressures to Establish Vulnerability Curves: Further Support for the Air-Seeding Hypothesis and Implications for Pressure-Volume Analysis”. Plant Physiology. 100 (1): 205–209. doi:1104/pp.100.1.205. PMC1075538. PMID 16652947.
- Schulte, P.J.; Hinckley, T.M. (1985). “A Comparison of Pressure-Volume Curve Data Analysis Techniques”. Journal of Experimental Botany. 36 (10): 1590–1602. doi:1093/jxb/36.10.1590.
- Cochard, Hervé; Forestier, Sébastien; Améglio, Thierry (2001-06-01). “A new validation of the Scholander pressure chamber technique based on stem diameter variations”. Journal of Experimental Botany. 52 (359): 1361–1365. doi:10.1093/jexbot/52.359.1361. ISSN0022-0957.