
Para que serve a hemoglobina em plantas?
Na planta, a atividade da enzima nitrogenase é mantida graças à expressão da leg-hemoglobina, uma proteína com elevada afinidade ao oxigênio, que sequestra o elemento do meio celular, ela está presente nos bacteriossomos em que a enzima nitrogenase estará ativa. O controle genético da expressão da leg-hemoglobina é função tanto do DNA nuclear da célula hospedeira quanto do DNA bacteriano. Além de eficiente, esse controle evidencia o relacionamento íntimo entre planta-fixador de nitrogênio ao longo da evolução.
Hoje sabe-se a importância existente entre o metabolismo do carbono e nitrogênio, o que tem sido foco de várias pesquisas para produtividade e controle ambiental, como a emissão de gás carbônico e efeito estufa na fixação do N.
A Leg-hemoglobina é composta por um grupo heme (como a hemoglobina presente no sangue dos mamíferos) ligado ao grupo protéico correspondente à globina encontrada no citosol intimamente associada aos simbiossomos. Essa proteína é importante para o funcionamento do sistema uma vez que é responsável pelo transporte e manutenção do nível de oxigênio a taxas que permitam, a um só tempo contemporizar o processo respiratório do bacterióide e o funcionamento da nitrogenase, enzima esta, bastante sensível ao oxigênio (a nitrogenase é oxidada pelo O2 tornando-se inativa).
Como é feito o controle da concentração de oxigênio ao redor da enzima nitrogenase?
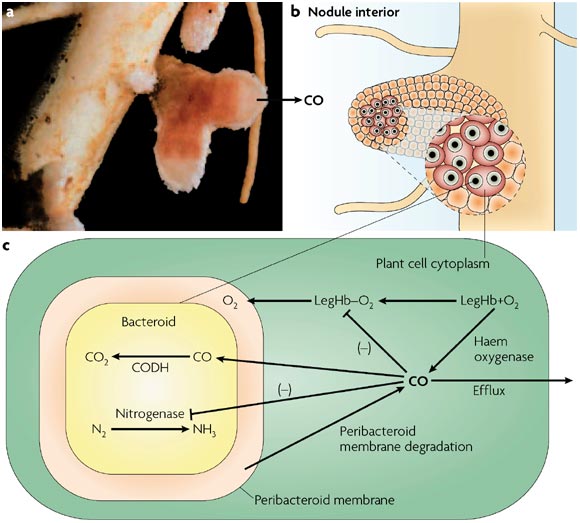
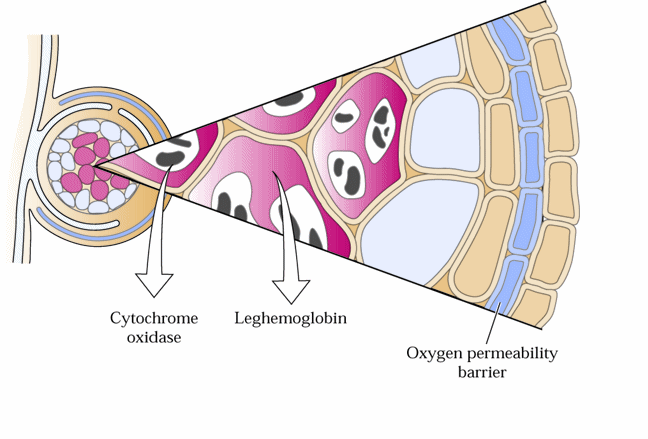
Os diferentes sistemas e organismos fixadores de nitrogênio apresentam mecanismos estruturais ou fisiológicos para manter baixa tensão de oxigênio ao redor da nitrogenase a fim de evitar sua inativação. Considerando os nódulos de leguminosos, a leg-hemoglobina controla, em parte, a disponibilidade de O2 junto ao bacterióide. No entanto, algumas características estruturais do nódulo, especialmente ligadas ao córtex como endoderme, fibras, esclereídes e inclusões de glicoproteínas nos espaços intercelulares, atuam como uma barreira à de fusão de gás, regulando o suprimento de oxigênio para o tecido infectado desempenhando um papel importante na proteção da nitrogenase. Adicionalmente no bacterióide, ocorre citocromo oxidase com alta afinidade pelo O2, auxiliando na respiração do bacterióide diante da baixa concentração desse gás. A figura abaixo apresenta resumidamente as três formas de controle de trocas gasosas no nódulo.
Em todas as leguminosas independentemente do tipo de nódulo, a fixação do nitrogênio não começa até que a planta possa dispor de parte dos produtos da fotossíntese que deverão ser canalizados para essa atividade, ou então, que haja um excesso de carbono (C) em relação ao nitrogênio (N) da planta. Em resumo ambos se beneficiam: nitrogênio fixado é fornecido para a planta e o carboidrato é fornecido para a bactéria. O mecanismo de regulação da atividade de fixação simbiótica seria a relação C/N interno.
A figura abaixo traz a troca de nutrientes entre a planta e o simbionte. Sabe-se que há um grande fluxo de nutrientes e gases entre o simbiossomo inserido na célula infectada e a célula não infectada. Análises proteômicas e transcriptômicas têm comprovado este grande fluxo pela presença de vários transportadores de açúcares (Suc/H+) entre as membranas. Também em feijão encontram-se transporte via difusão, mas a taxa de fluxo para o simbiossomo não é suficiente para suportar a fixação do nitrogênio. A planta fornece carbono para o simbiossomo principalmente na forma de malato. Primeiramente a sacarose é descarregada pelo floema nas células não infectadas, esta é clivada pela sacarose sintase (Susy) e invertase alcalina em hexose. Mutantes da sacarose sintase (Rug4) de Pisum sativum na qual perderam sua atividade enzimática não tiveram fixação de nitrogênio em seus nódulos comprovando que as enzimas são essenciais para a fixação. Também foram encontrados Susy e Invertase nas células infectadas, mostrando que a sacarose também pode ser transportada para estas células e ali serem hidrolisadas em hexoses. As hexoses são convertidas em Pep (fosfenolpiruvato) que podem ser carboxilados (Pepcase) a oxaloacetato e então reduzido (malato desidrogenase – MDH) a malato, para suprir o bacterióide em energia e esqueleto de carbono.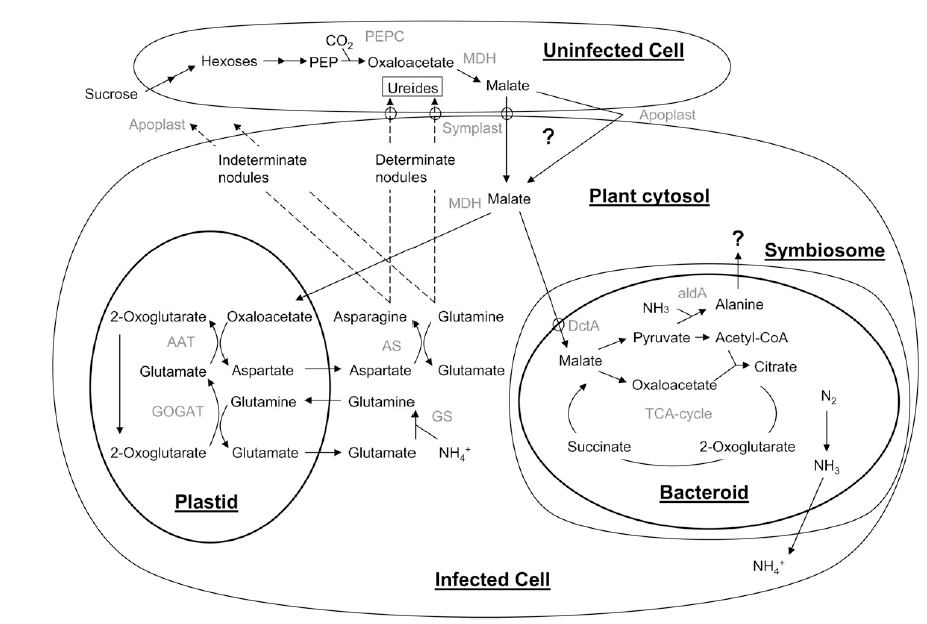
Em contrapartida o bacterióide fornece amônia pela fixação do N2 que é difundido pelo citosol e é assimilado pelas enzimas do metabolismo do nitrogênio (GS/GOGAT) formando aminoácidos. Outro caminho do bacterióide do fornecimento para a planta é a instantânea formação de alanina. Os aminoácidos formados tanto no bacterióide (alanina) como nas células infectadas, tomam caminho para a planta via xilema ou formam ureídeos sendo transportados.