
H+-ATPases (ATP fosfo-hidrolase) são proteínas integrais de membrana que utilizam a energia liberada na hidrólise do ATP para bombear prótons ativamente para o apoplasto ou para o vacúolo, de modo a controlar o pH citossólico. Dois tipos de ATPase’s, uma na plasmalema e uma outra no tonoplasto, e uma pirofosfatase inorgânica (utiliza a energia liberada na hidrólise do PPi) no tonoplasto, geram a força protomotriz para o transporte de solutos. Essas ATPase’s atuam no sentido de remover prótons do citossol, bombeando-os para o vacúolo ou para a parede celular. A operação dessas bombas extrusoras de prótons produz três efeitos importantes: aumento do pH no citossol, apesar de esse incremento ser de magnitude relativamente pequena, na medida em que há sistemas de tamponamento contra a perda de H+; redução do pH da parede celular (~5,5) e do vacúolo (4,5 a 5,9), contra valores de 7,3 a 7,6 no citossol, em razão da baixa capacidade tamponante daqueles compartimentos, e; o citossol torna-se eletronegativo em relação à parede e ao vacúolo, uma vez que ele libera H+, mas retém OH–. Em adição, há evidências da presença de ATPase’s que medeiam o contra-transporte de Ca2+/H+ (Ca2+ para a parede celular ou para o vacúolo e H+ para o citossol).
A operação das ATPase’s resulta, portanto, no estabelecimento de gradientes de µ e de pH. Apesar de esses gradientes não interferirem na absorção de moléculas neutras, eles desfavorecem grandemente a absorção de ânions. No entanto, o aumento na concentração protônica da parede celular (cerca de 100 vezes maior que a do citossol) favorece a reabsorção passiva de H+ no citossol, que ocorre a taxas muito baixas através da membrana plasmática, a menos que H+ se combine com um carregador ou mova-se através de um canal. Nesses carregadores, H+ geralmente é transportado para dentro da célula com um ânion acompanhante (co-transporte ousimporte). Nessa situação, H+ move-se a favor de seu gradiente de µ, enquanto o ânion é transportado ativamente. A estequiometria do co-transporte H+/ânion não está ainda estabelecida, mas acredita-se que mais de um próton seja transportado por cada carga negativa. A absorção passiva de H+ pode também ser usada para transportar, simultaneamente, cátions para o exterior celular (contra-transporte ou antiporte). Antiporte de Na+, por exemplo, é importante para as células radiculares eliminarem esse íon do citossol, especialmente quando a planta cresce em solos salinos.
No tonoplasto, a operação da H+-ATPase e da PPiase permite o armazenamento de íons e moléculas que podem ser recuperados pelo citossol quando necessário. Provavelmente, cátions são contra-transportados com H+, e ânions movem-se por uniporte, uma vez que o vacúolo é positivamente carregado em relação ao citossol. Há evidências também de que ânions possam ser contra-transportados com prótons nos vacúolos.
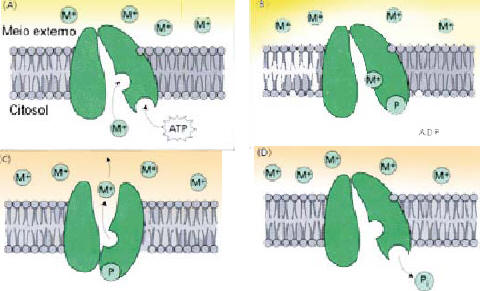
Figura 1: Mecanismo de funcionamento de uma bomba de íons. Esse tipo de proteína transportadora utiliza a energia diretamente acoplada a mesma, bombeando o íon contra um gradiente de concentraçãoEx: H+-ATPase, H+/Ca2+-AtPase e PPiase). (Motta, F. 2000)
Transporte Através de Carregadores e Canais
Carregadores e canais utilizam o gradiente de µ gerado pelas bombas extrusoras de prótons para transportar solutos passivamente através das membranas. Carregadores são proteínas integrais que ligam-se especificamente a um ou a muito poucos solutos relacionados. Provavelmente, sofrem uma mudança conformacional que facilita o transporte do soluto.
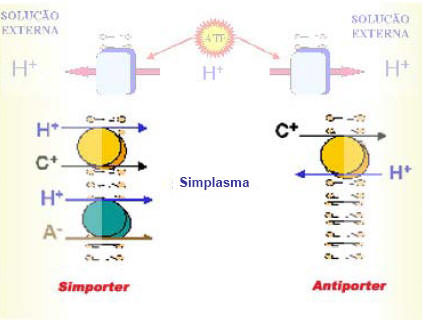
Figura 2 : Esquema do funcionamento de proteínas carregadoras, as quais utilizam o grandiente de prótons criado pelas bombas iônicas. Dois tipos de carregadores são apresentados: simporter, quando o transporte dos dois íons ocorre em uma mesma direção, ou antiporter, quando o transporte dos dois íons ocorre em direções opostas. Ex: triose/Pi, piruvato/ADP, sacarose/H ((Motta, F. 2000).
Canais são também proteínas integrais de membranas, coexistindo em duas conformações, “aberta” ou “fechada”, tendo, portanto, a faculdade de regular o fluxo de solutos, na medida em que a sua conformação é alterada. Canais permitem um rápido transporte passivo (uniporte), a taxas (106 a 108 íons por segundo) até cinco vezes maiores que aquelas observadas em carregadores. Até o presente, já foram identificados canais específicos para K+, Ca2+, H+ e Cl–, e postula-se a ocorrência de um canal para nitrato no tonoplasto. Entretanto, canais iônicos permanecem fechados na maior parte do tempo, e seu número por célula parece ser pequeno.
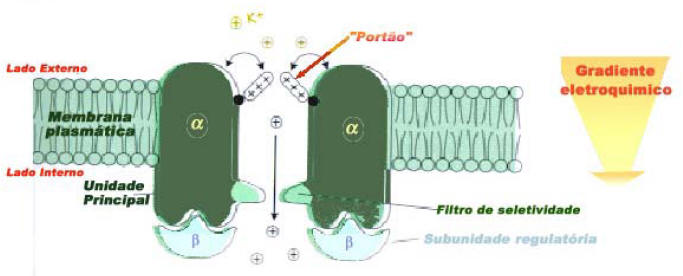
Figura 3: Estrutura de um canal de potássio, apresentando dois de seus peptídeos constituintes: suunidade alfa e beta. O portão do canal é aberto de acordo com um potencial elétrico de membrana, o qual é então o resultado também da ação de uma bomba iônica. Ex: canais de potássio, malato, e cloro (Motta, F. 2000).