
O floema é conhecido também como líber, constituindo uma camada entre a casca e a madeira, sendo responsável pelo transporte de assimilados da fotossíntese. Esse vaso condutor é formado por elementos crivados (células crivadas e elementos do tubo crivado), células companheiras ou albuminosas, fibras floemáticas e células de parênquima (Castro et al., 2009). Em plantas com crescimento primário, o floema também ocupa a porção externa dos caules, tendo como exceção as gramíneas, cujos vasos de floema e xilema estão distribuídos em vários feixes dispersos no córtex. Contudo, em cada feixe, o floema também ocupa a porção mais externa. (Castro, P.R.C; Kluge, R.A & Peres, L.E.P, 2005).
Os elementos crivados podem ser de dois tipos: células crivadas em gimnospermas e os elementos dos tubos crivados, que são células altamente especializadas de angiospermas. Os elementos do tubo crivado têm áreas crivadas especializadas, formando a placa crivada (Figura 1), que apresentam poros grandes circundados por calose. Eles são dispostos verticalmente, sendo conectados pelas placas crivadas formando tubos crivados e são acompanhados por células companheiras (Castro et al., 2009). Essa ligação entre elemento do tubo crivado e células companheiras é feita por um sistema de comunicação ramificado, chamado de plasmodesmos. Os plasmodesmos permitem a troca de substâncias entre as células.
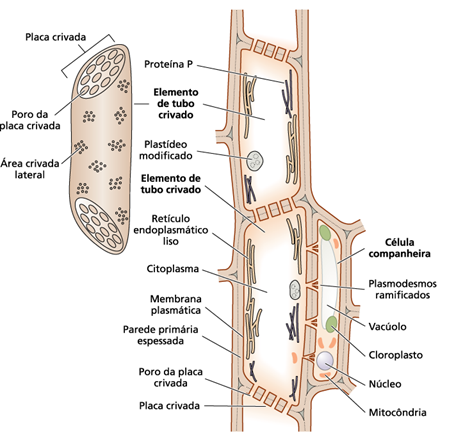
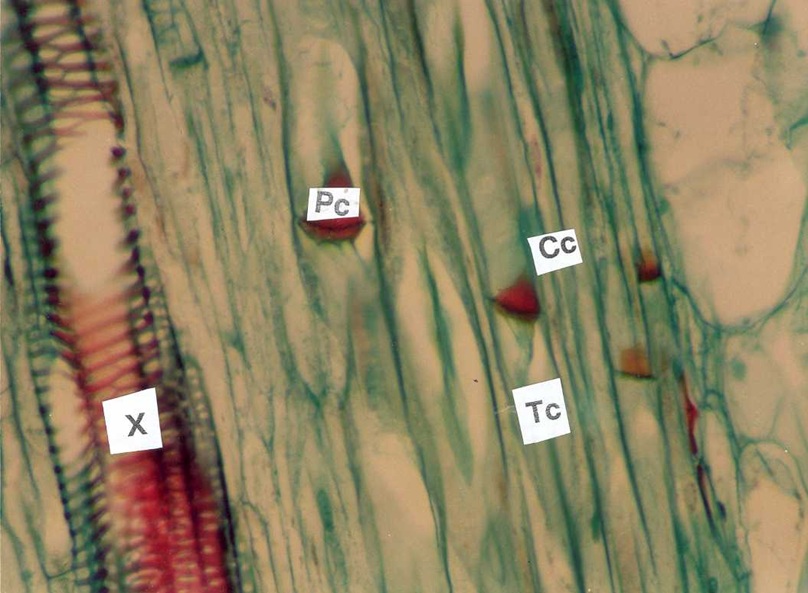
Figura 1: Placa Crivada. Seção longitudinal do caule de Curcubita pepo, mostrando os tubos crivados (TC) com as placas crivadas (Pc), as células companheiras (cc) e o xilema (x).
As células companheiras ao contrário dos elementos crivados são ricas em organelas, podendo sintetizar muitos compostos e substâncias como o ATP, que são transferidos para o elemento crivado, enquanto o elemento crivado tem função de realizar o transporte de assimilados da fotossíntese para o restante da planta (Figura 2). Algumas células companheiras alteraram a morfologia, formando invaginações da parede celular, sendo capazes de realizar transporte e então passaram a ser chamadas de células de transferência (ou intermediárias) (Kerbauy, 2004).
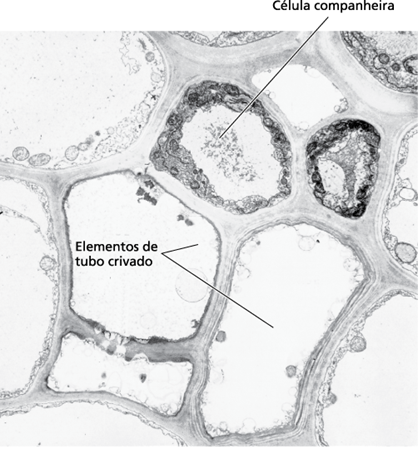
Figura 2: Células companheiras e elementos de tubo crivado
No floema a translocação não ocorre de forma ascendente ou descendente, e não é definida pela gravidade. O transporte ocorre da região produtora (fonte) para as regiões de metabolismo ou armazenamento (dreno) (Taiz & Zeiger, 2013). As fontes são denominadas áreas de produção de fotossintatos onde ocorre a translocação, para áreas de metabolismo intenso ou órgãos de reserva denominadas dreno. Fonte inclui qualquer órgão exportador, tipicamente uma folha adulta, completamente expandida, que é capaz de produzir fotossintato além de suas necessidades. Os maiores drenos durante a fase vegetativa são raízes e ápices caulinares enquanto na fase reprodutiva, os frutos se tornam dominantes. Nas nervuras terminais das folhas o carregamento do floema na região da fonte envolve o movimento dos produtos dos cloroplastos nas células do mesofilo para as células do tubo crivado (Zhang et al., 2005). Os órgãos fonte são responsáveis pela produção de assimilados a partir da fotossíntese e são representados principalmente pelas folhas. Os assimilados tanto podem ser usados como fonte energética necessária ao funcionamento da planta, através da respiração, como serem transportados e armazenados temporariamente em órgãos de reserva ou nos drenos, representados pelas raízes, meristemas e frutos das plantas. Segundo Marcelis (1996b), a força de fonte não é considerada neste processo, frequentemente, como exercendo efeito direto na distribuição de matéria seca, mas atua indiretamente, via formação de órgãos drenos.
Coll et al. (2001), acredita que entre os fatores que definem a força do dreno, o que mais se destaca é a proximidade, pois normalmente as fontes translocam nutrientes para os drenos que estão mais próximos delas, ou seja, translocam assimilados preferencialmente para drenos com os quais elas têm conexão vascular direta. Uma consequência prática a isso é que folhas que sombreiam outras mais próximas dos drenos de interesse devem ser eliminadas. Sendo um critério geral, as folhas superiores da planta normalmente translocam nutrientes para as folhas novas e caules em crescimento e as folhas da porção basal tendem a exportá-los para o sistema radicular. Já as intermediárias exportam seus produtos tanto para as partes superiores quanto para as inferiores. Durante o desenvolvimento a importância dos drenos é alterada, no crescimento vegetativo, os ápices caulinares e radiculares são os principais drenos, já no desenvolvimento reprodutivo os drenos são os frutos (Taiz & Zeiger, 2013).
As folhas responsáveis pela produção de compostos que serão transportados normalmente suprem os drenos com os quais possuem conexões vasculares diretas. Esses padrões de translocação podem ser alterados por ferimentos ou por erro de poda. Quando ocorre a eliminação de conexões vasculares, a rota de transporte passa a ser alternativa, sendo chamado de anastomose, forçando fontes mais distantes suprirem estes drenos. Como exemplo folhas inferiores suprindo as partes superiores e vice-versa, mas isso depende da espécie e órgão estudado (Taiz & Zeiger, 2013). Através do tamanho pode se determinar a força de um dreno (massa total do tecido) e atividade (taxa de importação e assimilados), mas também depende de outros fatores como suprimento de carboidratos, presença de outros drenos e resistência ao transporte ao longo do floema (Thorpe & Minchim, 1996).
O excedente em compostos fotoassimilados produzidos pela planta durante o período de atividade fotossintética, ficam imobilizados na forma de carboidratos insolúveis em órgãos aéreos e subterrâneos, sendo, então, mobilizados gradativamente em carboidratos solúveis durante o período de dormência. Essa mobilização é acelerada com o fim da dormência e os carboidratos solúveis conduzidos para as gemas em brotação que, por sua vez, formarão novos ramos e folhas. Posteriormente, as flores e os frutos são supridos, seguidos pelo câmbio, por novas gemas em formação e, finalmente, pelos tecidos que servem como depósito de carboidratos em órgãos subterrâneos e aéreos da planta (Baptist et al., 2009).
As substâncias para serem translocadas devem estar numa forma solúvel. Se forem insolúveis devem sofrer transformação de maneira a tornarem-se solúveis. O sentido da translocação é sempre da direção do órgão produtor ou de reserva (fonte) para o órgão consumidor (dreno), sempre que há demanda. A água e o carboidrato nela dissolvido a serem transportados, movem-se por fluxo de massa ao longo de um gradiente de pressão na direção do dreno de fotoassimilados. O que determina o volume do fluxo para um dado dreno é a distribuição diferencial de fotoassimilados dentro da planta, denominada partição (Tanaka & Fugita, 1979).
O caminho da translocação são os tecidos vasculares distribuídos por toda a planta, interligando as fontes aos drenos. O tecido vascular forma um sistema de tubos que pode direcionar, por partição, o fluxo de fotoassimilados para vários órgãos: folhas jovens, caule, raízes, frutos ou sementes. Quase sempre que há partição de fotoassimilados em beneficio de um determinado órgão e outro, possivelmente, ficará prejudicado, pois nem sempre a planta produz fotoassimilados o suficiente para abastecer todos os seus órgãos simultaneamente. Em feijoeiros, por exemplo, no início da floração, os fotoassimilados deixam de ser translocados para as raízes e tomam a direção das flores e frutos, onde a atividade metabólica passa a ser intensa com alta demanda por fotoassimilados (Tanaka & Fugita, 1979).
Desta maneira temos que o fluxo de fotoassimilados através do sistema vascular depende do estádio de desenvolvimento da planta. Plantas em crescimento intenso têm maior atividade metabólica e, portanto, maior demanda pelos seus diferentes órgãos e a taxa de fluxo é alta. Por outro lado, plantas maduras ou em crescimento lento, têm atividades metabólicas menores e, consequentemente, menores taxas de fluxo de fotoassimilados. Como resultados têm um menor crescimento radicular. Ao longo do ciclo das plantas nos diferentes eventos ontogenéticos, que ocorrem como consequências do seu desenvolvimento podem ocorrer mudanças no direcionamento dos fotoassimilados (Tanaka & Fugita, 1979).
A quantidade total de carbono fixada disponível para a folha é determinada pela taxa fotossintética, porém, a quantidade de carbono disponível para a translocação depende dos eventos metabólicos subsequentes. A regulação do direcionamento do carbono fixado para os vários caminhos metabólicos é chamada alocação (Taiz & Zeiger, 2013). Os fotoassimilados não utilizados pela folha são transportados para outros órgãos onde vão entrar no crescimento dos mesmos. Normalmente, o transporte da solução água e solutos na planta através do xilema ocorrem a favor de gradientes de concentração, sempre no sentido da maior para a menor concentração. No caso do transporte de solutos orgânicos, via floema, ocorre a favor de gradiente de pressão, independente do gradiente de concentração, ou de potencial da água (Tanaka & Fugita, 1979).
Substâncias transportadas no floema
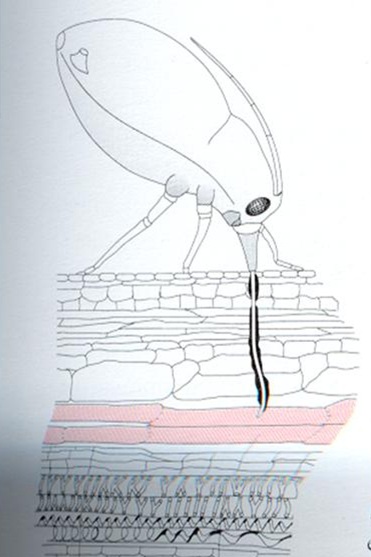
Muito do que sabemos hoje sobre transporte no floema é graças ao estudo dos afídeos (pulgões e cochonilhas). Esses insetos introduzem seu aparelho bucal (rostro), diretamente no tubo crivado e ao analisar o conteúdo sugado é deduzido o que é transportado no floema. Pulgões anestesiados com fumigação de CO2 têm seus rostros isolados e estes continuam exudando seiva, já que o floema está sobre pressão. Essas análises indicam que o principal soluto transportado no floema é a sacarose, sendo o constituinte onipresente de todos os conteúdos do floema. A concentração de sacarose transportada varia entre 0,3 a 0,9 M. Além da sacarose, em algumas famílias de plantas o floema também transloca outros açúcares não redutores (pois são menos reativos), tais como: rafinose (sacarose + galactose), estaquiose (sacarose + 2 galactoses) e verbascose (sacarose + 3 galactoses). Açúcares cujos grupos aldeído e cetonas foram reduzidos a álcool (manitol, sorbitol) também são translocados (Zimmermann & Ziegler, 1975 apud Lalonde et al, 2003).


A substância transportada em maior quantidade pelo floema é a água, sendo que os solutos transportados estão dissolvidos nela. O transporte de aminoácidos e amidas (fontes de nitrogênio) é consideravelmente baixo quando comparado com o transporte de carboidratos. Os aminoácidos e amidas são encontrados no floema geralmente na forma de glutamato e aspartato e suas respectivas amidas glutamina e asparagina, mas nunca na forma de nitrato (Taiz & Zeiger, 2013; Kerbauy, 2004). O nitrogênio é transportado na forma orgânica e inorgânica. No floema o transporte é preferencialmente na forma orgânica, já no xilema ele pode ser transportado como nitrato ou como parte de molécula orgânica (Taiz & Zeiger, 2013).
O floema também transporta substâncias sinalizadoras, sendo importante na comunicação entre as várias partes das plantas. Entre as substâncias sinalizadoras transportadas no floema, estão os hormônios vegetais (auxina, citocinina, giberelina, ácido abscísico). Outro importante sinal transportado pelo floema é o RNA mensageiro (mRNA). Em experimentos de enxertia combinando uma planta de genótipo mutante específico com outra de genótipo normal, pode se provar que substâncias passadas pelo floema são capazes de modificar o fenótipo de plantas não mutantes. A substância em questão é o mRNA da planta mutante (Kim et al., 2001). De modo semelhante, é bem conhecido que ao enxertarmos plantas não induzidas para o florescimento em plantas induzidas, uma substância desconhecida (florígeno) é capaz de ser passada através do floema para a planta não induzida, provocando seu florescimento (Aukerman & Amasino, 1998). Além de RNA, o floema também transporta outras macromoléculas como peptídeos (ex. sistemina) e partículas virais (Oparka & Santa Cruz, 2000). Nucleotídeos fosfato e proteínas também são encontrados na seiva elaborada no floema. As proteínas que são encontradas são as P, responsáveis pela obstrução do vaso em caso de danificação, proteínas quinases, tiorredoxina, ubiquitina (degradação de proteínas), as chaperonas (dobramento de proteínas), inibidores de proteases (Taiz & Zeiger, 2013; Kerbauy, 2004).
Apesar do floema não transportar nitrato, ele transporta muitos nutrientes minerais. Os solutos inorgânicos que se movem no floema são potássio, magnésio, fosfato e cloreto (Taiz & Zeiger, 2013; Kerbauy, 2004). O potássio é o nutriente presente em maior quantidade no floema, seguido de fósforo, magnésio e enxofre. O potássio, juntamente com a sacarose, é o principal componente osmótico da seiva do floema (Kerbauy, 2004).
A taxa de movimentação pelos elementos crivados no floema pode ser medida pela velocidade e pela taxa de transferência da massa. A velocidade está relacionada com a distância linear percorrida por unidade de tempo, e a taxa de transferência de massa é quantidade de material que passa pelo floema e elementos do tubo crivado por unidade de tempo. Geralmente a velocidade de translocação no floema é alta, variando de 0,3 a 1,5 m h-1 (30 a 150 cm por h-1). Dessa forma a difusão não pode ser responsável pelo processo por ser um mecanismo muito lento, sendo aproximadamente 1m em 32 anos, assim é necessário um movimento por fluxo de pressão (Taiz & Zeiger, 2013). Esse modelo de transporte foi proposto por Ernst Münch em 1930, sendo o fluxo ocasionado por gradiente de pressão gerado entre a fonte e o dreno (delta ψp). Esse gradiente ocorre por consequência do carregamento e descarregamento do floema. Para que a translocação ocorra é necessário gasto de energia tanto na fonte quanto no dreno. Nas fontes é necessário energia para transportar os produtos da fotossíntese para os elementos crivados, o que é chamado de carregamento do floema. Nos drenos a energia é usada para transportar os compostos do elemento crivado para a célula-dreno, o que é chamado de descarregamento do floema (Taiz & Zeiger, 2013).
Veja também: